
Птерозавры, гигантские летающие рептилии мезозойской эры, доминировали в небесах нашей планеты по меньшей мере 150 миллионов лет (с позднего триаса до конца мела). Множество особенностей — от облегченного скелета до специфического строения конечностей — указывают на глубокие адаптации к воздушному образу жизни. Однако, как показывают многочисленные следы, сохранившиеся по всему земному шару, с определенного времени эти рептилии начали активно осваивать и наземное перемещение, чувствуя себя на четырех конечностях не менее комфортно, чем пролетая на широко распахнутых крыльях над головами динозавров. В новом исследовании, опубликованном в журнале Current Biology, международный коллектив исследователей смог связать окаменелые следы с тремя группами птерозавров, которые их оставили. Судя по полученным результатам, птерозавры начали адаптироваться к передвижению на четырех лапах еще со средней юры, и к позднему мелу часть из них превратилась в полуназемных хищников, активно ищущих корм на суше.
Несомненно, главенствующими летающими позвоночными мезозоя были птерозавры, отличавшиеся огромным разнообразием форм и размеров. Поскольку анатомически и, очевидно, физиологически эти животные были глубоко адаптированы к летающему образу жизни, палеонтологи в первую очередь уделяли внимание их летным качествам (см., например, Полет птерозавров в ходе эволюции неуклонно совершенствовался, «Элементы», 02.11.2020; Крылатые гиганты прошлого были неспособны к динамическому парению, «Элементы», 27.05.2022), считая способности к наземному передвижению невысокими и связывая их образ жизни в первую очередь с морскими экосистемами (P.
Wellnhofer, 1987. Terrestrial locomotion in pterosaurs).Однако на рубеже тысячелетий начали накапливаться свидетельства в пользу того, что птерозавры много и охотно передвигались по земле, забираясь вглубь суши и оставляя там свои характерные отпечатки (D. Unwin, 1996. Pterosaur tracks and the terrestrial ability of pterosaurs; J. Mazin et al., 2003. Ichnological evidence for quadrupedal locomotion in pterodactyloid pterosaurs: trackways from the Late Jurassic of Crayssac (southwestern France); M. Lockley et al., 2008. The fossil trackway Pteraichnus is pterosaurian, not crocodilian: Implications for the global distribution of pterosaur tracks). Также последовали переоценки морфологии скелета, разнообразия питания и летных качеств, что показало, насколько широкий спектр экологических ниш занимали крылатые рептилии на мезозойской Земле (M. Witton, D. Naish, 2008. A Reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology; J. Bestwick et al., 2020. Dietary diversity and evolution of the earliest flying vertebrates revealed by dental microwear texture analysis; M. Witton, M. Habib, 2010. On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness). По-видимому, упор на развитие полетных характеристик не ограничил совершенствование альтернативных способов перемещения, и наравне с динозаврами в сухопутных экосистемах птерозавры выполняли роль как мелких хищников, так и добычи.

За последние тридцать лет было найдено множество новых образцов следов птерозавров, причем в некоторых местах их явно больше, чем ископаемых костей (J. Jung, M. Huh, 2024. New Pterosaur Tracks from the Hwasun Seoyuri Tracksite (Turonian) of South Korea: Implications for their Ecological Niche and Habitat). Это порядком затрудняет определение принадлежности следов хотя бы до уровня семейства (см. картинку дня Следы невиданных зверей), и в результате даже активное накопление ископаемого материала дает не так много сведений о биологии таксонов. Обнаруживая следы крылатых ящеров, ученые приходят к выводу лишь о присутствии неопределенных птерозавров в регионе в какой-то период времени, но не используют эти сведения при изучении биогеографии или макроэволюции конкретных клад (Y. Yu et al., 2023. Complex macroevolution of pterosaurs).
Конечно, на то есть веские причины, и в первую очередь это сложности с отнесением нового ихнотаксона к конкретной кладе птерозавров. Чаще всего ископаемые отпечатки относят к продвинутым птерозаврам-птеродактилоидам (В Аргентине обнаружили переходное звено между примитивными и прогрессивными птерозаврами, «Элементы», 13.12.2024), хотя этот подход не общепринятый и может потребовать пересмотра (M. Witton, 2015. Were early pterosaurs inept terrestrial locomotors?). Редкие попытки отнести отпечатки к конкретной группе основываются на косвенных доказательствах, таких как абсолютный размер следов или их стратиграфический возраст (K. Hwang et al., 2002. New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, southwestern Korea), либо же на ассоциации следов с костными остатками, которые и считаются «следообразователем» (Y. Li et al., 2025. First deciphering of large pterosaur footprints and their trackmaker in the Junggar Basin, China).

В новом исследовании ученые из Великобритании и США использовали 3D-моделирование, подробный количественный анализ и сравнение следов с костными остатками, чтобы выделить ключевые синапоморфии трех крупных групп птерозавров. Работа была проведена на 52 образцах, обнаруженных на всех континентах за исключением Австралии и Антарктиды; возраст отпечатков составлял от 161 до 66 миллионов лет.
В первую очередь палеонтологи сосредоточились на отпечатках стоп, поскольку отпечатки передних конечностей птерозавров сравнительно консервативны по форме и несут меньше таксономически ценной информации. Учитывая, что все сохранившиеся отпечатки передних конечностей отличаются чрезмерно длинным третьим пальцем, авторы пришли к выводу, что отпечатков таксонов с более пропорциональными пальцами (например, анурогнатид; см. картинку дня Насекомоядный птерозавр) пока найдено не было.
Судя по сохранившемуся образцу птерозавра с мягкими тканями, у этих ящеров были «тощие лапки» с минимальным количеством мягких тканей вокруг ступни и пальцев, что характерно и для современных летающих позвоночных — птиц и рукокрылых (E. Frey et al., 2003. A new species of tapejarid pterosaur with soft-tissue head crest). Это облегчает работу ученых, которым приходится закладывать меньшие поправки на разницу между скелетной основой и следом живого птерозавра, и повышает точность измерений. Проанализировав все имеющиеся следы, исследователи отнесли их к поздним птерозаврам-птеродактилоидам, а также выделили три морфотипа следов, каждый из которых соотносится с конкретной кладой птеродактилоидов.

Морфотип I был отнесен к археоптеродактилоидам (Archaeopterodactyloidea), или ктенохазматоидам — ранним птеродактилоидам, существовавшим с конца юрского по ранний меловой периоды. Их остатки обнаружены в Африке, Евразии и Северной Америке. Многие виды этой клады отличались необычным строением черепа и зубов, в частности, именно к ктенохазматоидам относится птеродаустро (Pterodaustro guinazui; см. картинку дня Птеродаустро и его челюсть-щетка). Для морфотипа I характерны сравнительно крупные отпечатки стоп почти треугольной формы, длинные плюсневые кости и короткие пальцы. Первоначально эти следы считали принадлежащими крокодиломорфу, но затем была доказана их принадлежность птерозавру (K. Padian, P. Olsen, 1984. The Fossil Trackway Pteraichnus: Not Pterosaurian, but Crocodilian; M. Lockley et al., 2008. The fossil trackway Pteraichnus is pterosaurian, not crocodilian: Implications for the global distribution of pterosaur tracks). От отпечатков других птерозавров отпечатки морфотипа I отличаются соотношением длин плюсневых костей: первая длиннее второй, вторая длиннее третьей, третья длиннее четвертой. Такое распределение крайне нетипично для диапсидных рептилий, включая крокодиломорфов, у которых первая плюсневая кость сравнительно короткая.
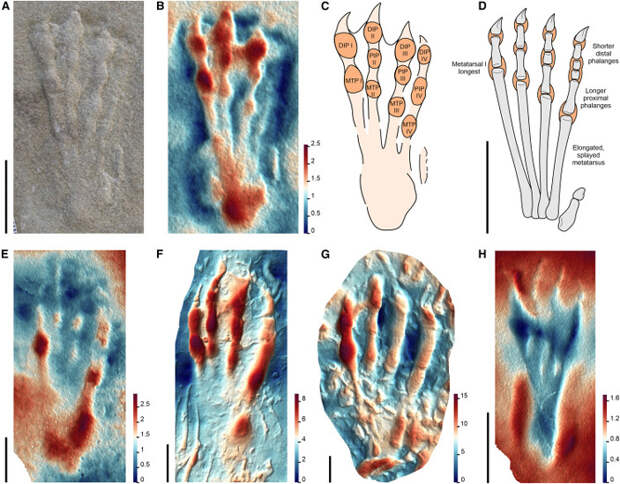
Морфотип II отнесли к джунгариптеридам (Dsungaripteridae), семейству птерозавров из раннего мела Евразии и Южной Америки. В данном исследовании рассматривались два ихнотаксона из раннего мела Китая, Pteraichnus wuerhoensis и Pt. junggarensis. Эти следы отличались от следов морфотипа I относительными длинами плюсневых костей (кость I примерно равна кости II, и обе сравнительно короче, чем в отпечатках ктенохазматоидов) и костей пальцев (первый палец укорочен, остальные три примерно равны по длине, в то время как у морфотипа I все четыре пальца примерно одинаковы, см. рис. 2). Такое соотношение костей пальцев характерно для джунгариптерид Dsungaripterus weii и Noripterus complicidens и птеранодонов Pteranodon, однако пропорции плюсневых костей у последних отличаются, что позволяет отнести отпечатки морфотипа II именно к джунгариптеридам. Впрочем, поскольку результаты анализа основаны на крайне ограниченной выборке (четыре отпечатка и одна полностью сохранившаяся стопа джунгариптерида), воспринимать их следует с некоторой осторожностью.

Наконец, следы морфотипа III были наиболее распространены географически и были отнесены к неоаждархиям (Neoazhdarchia), подгруппе аждархоидов (Azhdarchoidea), крупных меловых птерозавров со сравнительно короткими крыльями. Следы такого морфотипа отличались узкой удлиненной стопой со слабыми отпечатками пальцев и хорошо отпечатавшейся пяткой, на которой виден след пяточной подушечки — явной адаптации к наземному передвижению. Соотношение плюсневых костей у следов такого морфотипа отличается от предыдущих (плюсневая кость I примерно равна по длине плюсневой кости II или короче ее), и все плюсневые кости тесно сжаты. Поскольку хорошо сохранившиеся отпечатки такого морфотипа — большая редкость, а развитые подушечки затрудняют оценку длин плюсневых костей, авторы рекомендуют при определении принадлежности таких следов использовать явную синапоморфию — хорошо отпечатывающуюся пятку с развитой подушечкой.
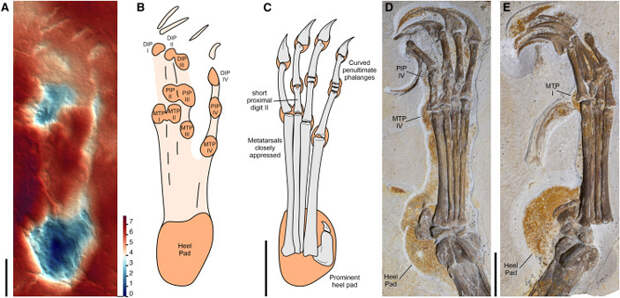
В настоящее время наши знания о следах птерозавров все еще очень ограничены: к примеру, к ихнотаксону Pteraichnus относятся следы, которые в данном исследовании отнесены к разным морфотипам и, соответственно, кладам птерозавров. Используя примененную авторами методологию, можно уточнять время существования, экологию и особенности поведения представителей конкретных клад, расширяя наши знания о биологии птерозавров.
Несмотря на относительно многочисленную выборку, ни один из изученных отпечатков не был с уверенностью отнесен к ранним птерозаврам-рамфоринхоидам (Rhamphorhynchoidea), для которых характерен хорошо развитый пятый палец стопы, сопоставимый по размерам с остальными. Все задокументированные следы относятся к временному периоду от средней юры до позднего мела (а появились первые птерозавры еще в триасе), причем костные остатки первых птеродактилоидов несколько моложе древнейших следов морфотипа I (C. Zhou et al., 2017. Earliest filter-feeding pterosaur from the Jurassic of China and ecological evolution of Pterodactyloidea). Впрочем, это соответствует результатам филогенетических исследований, предполагающих появление птеродактилоидов в среднем юрском периоде.
Можно предположить, что рамфоринхоиды в основном лазали по деревьям и скалам, не спускаясь, к примеру, на полосы прибрежной грязи и ила, в которых часто сохраняются ископаемые отпечатки, тогда как у первых птеродактилоидов развились стопы и кисти, лучше приспособленные для передвижения по земле, — и в ископаемой летописи появились первые следы птерозавров (R. Smyth et al., 2024. Hand and foot morphology maps invasion of terrestrial environments by pterosaurs in the mid-Mesozoic). Стоит, правда, отметить, что не все птеродактилоиды были хорошими ходоками: у представителей некоторых групп, например, аньянгуэрид (Anhangueridae), были редуцированные кости таза, сравнительно короткие ноги и маленькие ступни, так что вряд ли они часто оставляли свои отпечатки в палеонтологической летописи.
Судя по распределению следов, ктенохазматоиды были неотъемлемой частью прибрежных морских экосистем и, судя по обилию отпечатков, в свое время буквально кишели на отмелях, отфильтровывая из воды мелкую живность или охотясь за рыбешкой. Джунгариптериды, в свою очередь, тяготели к мелководным рекам и озерам, где, судя по их характерным челюстям с беззубыми кончиками и крепкими зубами, искали моллюсков с твердыми панцирями. Наконец, следы неоаждархий обнаруживаются исключительно в континентальных отложениях до самого конца мелового периода, после чего они начинают попадаться в прибрежных регионах. Любопытно, что в тех местах, где следы неоаждархий сохраняются вместе со следами сухопутных позвоночных, направление следовых дорожек птерозавров совпадает с дорожками динозавров, которые, натолкнувшись на водоем, воспринимали его как препятствие и обходили по краю — то есть, даже натолкнувшись на преграду, птерозавры не поднимались на крыло, а предпочитали обойти ее пешком (L. Xing et al., 2015. Reanalysis of Wupus agilis (Early Cretaceous) of Chongqing, China as a Large Avian Trace: Differentiating between Large Bird and Small Non-Avian Theropod Tracks).
Подводя итоги, с помощью количественного анализа и выделения ключевых синапоморфий палеонтологи теперь смогут определять следы птерозавров не до уровня отряда и подотряда, но до более узкой клады, что позволит больше узнать об их экологии, а также географическом и временном распределении. Судя по полученным данным, птерозавры начали активно вторгаться в наземные экосистемы уже в среднем юрском периоде, адаптируясь к передвижению по суше, что в конечном итоге привело к появлению «цапель-монстров» позднего мелового периода (см. картинку дня Крылатые чудовища древней Трансильвании), наводивших ужас на мелких динозавров и других сухопутных позвоночных по всему земному шару.
Конечно, ни одно летающее позвоночное животное не проводит жизнь исключительно в воздухе, спускаясь на землю хотя бы для размножения. Возможно, ранние птерозавры, подобно современным мелким воробьиным вроде чижей или клестов, в основном отдыхали и размножались на скалах и ветвях деревьев, откуда им было удобно взлетать, спрыгивая прямо в воздух: об этом свидетельствуют их цепкие кисти с крупными когтями и относительно короткие стопы с длинными хватательными пальцами. Поздние птерозавры начали больше времени проводить на земле, как нелетающие четвероногие, чему способствовали более длинные ноги, редукция хвостовой перепонки, удлиненные и крепкие кисти и стопы: подобно куликам или голубям, они могли искать пищу на земле, поднимаясь на крыло только в случае опасности или при необходимости добраться до удаленного места (R. Smyth et al., 2024. Hand and foot morphology maps invasion of terrestrial environments by pterosaurs in the mid-Mesozoic).
Наконец, стоит отметить, что активно передвигаться по суше птерозавры могли начать еще в раннем мезозое. К примеру, британский палеонтолог Марк Уиттон (Mark Witton) отмечал, что отсутствие следов птерозавров с триаса по среднюю юру может и не отражать их привычки в передвижении (M. Witton, 2015. Were early pterosaurs inept terrestrial locomotors?): остатков крылатых рептилий из этого временного периода в принципе меньше, чем многих современных им четвероногих, что может быть следствием особенностей экологии (например, птерозавры в основном заселяли леса и горы, где остатки сохраняются плохо) или биогеографии (на ранних этапах своей эволюционной истории птерозавры могли быть распространены нешироко). Вдобавок, ранние птерозавры были в основном некрупными животными размером от галки до орла, так что их следы были невелики и неглубоки: возможно, мы их просто еще не нашли, а если и нашли, то не распознали?
В таком случае обилие следов птеродактилоидов во второй половине мезозоя — это не только следствие их анатомической приспособленности к наземному передвижению, но и результат активного освоения прибрежных и речно-озерных экосистем, где они искали корм подобно современным куликам. Рамфоринхоиды же — в основном насекомоядные и рыбоядные виды, искавшие добычу в открытом море, — с меньшей вероятностью стали бы бродить вдоль линии прибоя или по берегу озера, оставляя следы в иле или мягкой грязи. Так что, удивляясь отсутствию их ископаемых следов или объясняя это исключительно анатомическими ограничениями, мы удивляемся отсутствию следов двугорбого верблюда в песчаных дюнах или на дне высохшей австралийской реки.
Источник: R. S. H. Smyth, B. H. Breithaupt, R. J. Butler, P. L. Falkingham, D. M. Unwin. Identifying pterosaur trackmakers provides critical insights into mid-Mesozoic ground invasion // Current Biology. 2025. DOI: 10.1016/j.cub.2025.04.017.
Анна Новиковская
Свежие комментарии