
Выход на сушу и — как следствие — освоение множества новых экологических ниш, позволил тетраподам (четвероногим позвоночным) осуществить масштабную эволюционную радиацию: во много раз возросло количество их видов, а вместе с ним и высших таксонов. Отчасти рост разнообразия и разделение эволюционных путей различных групп (появление амфибий, рептилий и предков млекопитающих) связывают с изменением механизма питания. Вышедшим на сушу позвоночным пришлось перейти от питания через всасывание добычи вместе с водой к принципиально новым способам добычи, обработки и проглатывания пищи. Это, в свою очередь, требовало соответствующих адаптаций в строении ротового аппарата. Однако палеонтологические данные показывают, что выход на сушу и изменение строения челюстей не совпадают хронологически: освоение суши состоялось в девоне и раннем каменноугольном периоде, а челюсти стали меняться лишь на рубеже каменноугольного и пермского периодов. Ученые из Бристольского университета задались вопросом, что могло вызвать такую рассинхронизацию. Их ответ — челюсти позвоночных начали меняться не из-за непосредственного выхода на сушу, а только после появления и распространения растительноядности.
При переходе из воды на сушу позвоночным пришлось осваивать новые механизмы питания. До выхода на сушу многие позвоночные добывали пропитание, засасывая воду в рот вместе с добычей, многие из них были хищниками-засадчиками. На суше таким способом охотиться нельзя, поэтому первым наземным позвоночным приходилось учиться добывать пропитание другими методами. Логично было бы предположить, что такие радикальные изменения в экологии и поведении наземных первопроходцев должны были повлечь за собой и морфологические изменения, а существа со «старым» типом питания, — а значит, вероятно, и челюстей — отсеялись бы отбором. Однако ранние наземные позвоночные на протяжении десятков миллионов лет сохраняли старый — «водный» — тип челюстей: практически параллельное их расположение в открытом состоянии, низкое положение подъязычной кости. И успешно использовали его для пропитания. Ситуация стала меняться лишь с появлением растительноядных животных. Авторы обсуждаемой статьи поставили перед собой задачу построить с помощью большого объема данных хронологию эволюции челюстей, а также количественно оценить, могло ли освоение растительноядности повлиять на изменения в строении черепа наземных позвоночных.
Согласно скомпилированным ими данным, с момента выхода позвоночных на сушу в конце девонского периода (~360 миллионов лет назад) вплоть до примерно 265 миллионов лет назад челюсти позвоночных практически не меняли строение. Были лишь несколько очень локальных (по времени) всплесков разнообразия. Только в конце каменноугольного и начале пермского периода начинают возникать новые варианты строения челюстей, радикально новые по сравнению с предками тетрапод.
У древних лопастеперых рыб и первых амфибий челюсти часто напоминали по своему строению капкан — широкие и плоские, с рядом острых зубов, приспособленные к коротким быстрым движениям, во время которых они целиком засасывали добычу с потоком воды. Авторы статьи предполагают, что челюсти ранних наземных позвоночных, хотя и имели немного другую функцию (использовались для хватания) могли выполнять ее со схожим строением, а потому долго не менялись и занимали морфопространство (см. morphospace и статью А. Маркова Как измерить морфологическое пространство?), не отличимое статистически от челюстей древних лопастеперых.
Морфологическое пространство (или просто морфопространство) — математическое понятие, позволяющее количественно оценивать морфологические различия между организмами. Это многомерное пространство, в котором каждая координата соответствует своему количественному морфологическому признаку, будь то высота, ширина, толщина или что-то еще. Многомерные данные с помощью метода главных компонент можно свести к 2- или 3-мерному, что позволяет выявлять статистически значимые отличия или схожести в строении изучаемых органов. Кроме того, по положению в морфопространстве можно предсказывать экологические черты организма.
Авторы проверяли гипотезу о том, что именно освоение растительноядности стало движущей силе развития амниот (рептилий, включая птиц, и синапсид, включая млекопитающих). Делали это, сравнивая положение челюстей различных тетрапод в морфологическом пространстве. Авторы изучили челюсти 137 видов четвероногих позвоночных и лопастеперых рыб (начиная с девона и заканчивая пермью) по различным характеристикам, а также построили гипотетические челюсти с невозможной геометрией, чтобы установить, какие факторы лимитируют эволюцию челюстей в том или ином направлении.
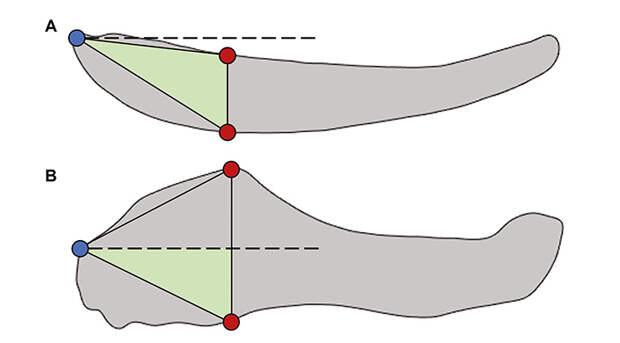
Исследователи измерили челюсти по следующим характеристикам:
1) Сила челюсти. Нагрузка, испытываемая челюстью при укусе, — один из факторов отбора, который может приводить к изменению морфологии челюсти. В частности, из-за него могут меняться положение челюстного сустава и мышц, управляющих челюстью.
2) Эффективность вращения — скорость перемещения кончика челюсти относительно челюстного сустава.
3) Механический выигрыш — отношение силы, приложенной мышцами челюсти, к силе, создаваемой челюстью. Более статичные типы питания (долгое разжевывание пищи с закрытым ртом) приводят к увеличению силы, создаваемой закрытой челюстью, в то время как динамичные (быстрые, короткие и малочисленные укусы) — к увеличению скорости движения челюсти из открытой позиции, для чего необходимо по-разному ориентировать челюсть и мышцы, приводящие ее в движение. У амниот, благодаря их более высоким, чем у анамний, черепам и более глубоким флангам крыловидного отростка, более эффективный для статичного питания ротовой аппарат, что, вероятно, предрасполагает их к развитию растительноядности.
4) Высота челюсти — высокие челюсти создают излишнее трение при плавании, и потому, вероятно, водным позвоночным зачастую невыгодно иметь высокий, объемный череп.
Благодаря проведенным измерениям, авторы смогли протестировать два предположения. Первое состоит в том, что морфопространство, которое делят между собой водные и наземные хищники, оптимально как для наземных, так и для водных животных. Второе — что морфопространство, занимаемое челюстями травоядных, оптимально для амниот, но не для анамний. Иными словами, авторы предполагают, что для того, чтобы продолжать быть хищниками, наземным позвоночным не надо было менять свой челюстной аппарат, в то время как переход на растительное питание запустил отбор на новое строение челюстей, которое стало «фишкой» амниот и позволило им создать уникальное видовое и морфологическое разнообразие.
Расположив челюсти исследуемых позвоночных на временной линии, можно выделить три основных этапа в эволюции позвоночных. Первый — середина и конец девона, когда происходило формирование наземных групп позвоночных. Второй — начало и середина каменноугольного периода, когда происходило первое значимое увеличение разнообразия в строении челюстей, этот же этап соответствует появлению в палеонтологической летописи растительноядных позвоночных и амниот, однако к его концу увеличение разнообразия затормаживается. Третий этап — пик в разнообразии челюстей в середине перми и стабильный уровень разнообразия в строении челюстей вплоть до пермо-триасового вымирания, самого масштабного вымирания за историю Земли. При этом пик разнообразия наблюдается в основном среди наземных растительноядных, в то время как хищники и водные позвоночные сохранили схожие уровни разнообразия челюстей.

Рисунок 2 демонстрирует, что морфологическое пространство амниот и анамний значительно пересекается, однако у анамний меньше разнообразия в строении челюстей, в то время как древние амниоты чаще имеют более экстремальные, узкоспециализированные типы челюстей. При этом большинство экстремальных вариантов строения обнаруживается именно у травоядных амниот, имеющих более глубокие и обширные челюсти. Челюсти хищников более сходны между собой по строению. Анализ оптимизации реальных и гипотетических челюстей показал, что растительноядность коррелирует с более глубокими челюстями, с изменением их изгиба с выпуклого на вогнутый и снижением положения челюстного сустава.
Достижение гипотетических крайне экстремальных форм челюсти (например, сочетающих в себе крайне высокие или крайне низкие значения по всем параметрам — силе, высоте, механическому выигрышу и эффективности вращения или очень вогнутые или выпуклые челюсти) невозможно не только из-за того, что это может нарушить их функцию, но и из-за косвенных факторов, например, эволюционной истории группы или влияния на челюсти нефункциональных признаков. В случае исследуемых групп позвоночных, в частности, достигнуть экстремальных форм челюсти было невозможно из-за необходимости компромисса между различными параметрами, поскольку одними признаками приходится жертвовать ради усиления других. Ограничения, наложенные на эволюцию челюстей, могут также объяснить, почему между тремя периодами эволюционных изменений есть стазисы.
Авторы предполагают, что достигнутый в водной среде компромисс между характеристиками челюстей ограничивал их развитие в наземной среде, поскольку такое строение не было под нее оптимизировано. В частности, строение мышц, управляющих челюстью, делало невозможным увеличение ее высоты, поскольку именно низкая челюсть оптимальна для мышечного строения у водных и ранних наземных хищников. Водный тип челюстей оптимизирован для минимизации стресса и высоты челюсти, а также для максимизации вращательной эффективности и механического выигрыша. Под те же показатели оптимизированы и челюсти наземных хищников, в отличие от наземных растительноядных позвоночных.
Тем не менее, по данным авторов, одного высвобождения челюстей от данных ограничений было недостаточно: согласно их расчетам, освоение растительноядности должны были спровоцировать и сторонние факторы. В позднем палеозое разнообразие типов челюстей продолжало расти, что, с одной стороны, было вызвано тем, что челюсти продолжили меняться и начали занимать неизведанные до этого регионы морфопространства, а с другой — тем, что в пермском периоде во время кунгурско-роудского вымирания (примерно 275 миллионов лет назад) исчезли многие таксоны с промежуточным строением челюстей, что увеличило долю высокоприспособленных и уникальных типов строения в выборке и повлияло на результаты статистического анализа.
Результаты, полученные авторами, раскрывают хронологию эволюции черепов ранних наземных позвоночных, а также указывают на то, что изменения в строении челюстей наземных позвоночных, а также рост разнообразия в их строении совпали с появлением и распространением растительного питания у наземных четвероногих и с появлением амниот. Благодаря количественным данным ученые смогли подтвердить, что водный тип строения челюстей был оптимален не только для водного образа жизни, но и для наземных хищников. Их результаты также показали, что освоение растительноядности было одним из движущих факторов в увеличении разнообразия строения черепов позвоночных. Оно сопровождалось не только изменением строения черепа, но и, например, изменениями строения мышц челюстей, кишечника и нефункциональных (потерявших свою изначальную функцию, однако присутствующих в организме и влияющих на другие характеристики) признаков. Суммарно, эти изменения в дальнейшем привели к увеличению разнообразия позвоночных, освоению ими большого количества экологических ниш и невероятному эволюционному успеху.
Источник: Harry O. Berks, Pablo S. Milla Carmona, Philip C. J. Donoghue, Emily J. Rayfield. The evolution of herbivory, not terrestrialisation, drove morphological change in the mandibles of Palaeozoic tetrapods // Evolutionary Journal of the Linnean Society. 2025.DOI: 10.1093/evolinnean/kzaf004.
Владимир Вержбицкий
Свежие комментарии