Изучение ДНК из осадков мангровых болот и эстуариев Китая позволило обнаружить 136 новых видов асгардархей — группы, к которой принадлежали предки эукариот. В результате удалось уточнить положение эукариот на эволюционном дереве асгардархей, а также реконструировать тип метаболизма предков эукариот. Хотя те два вида асгардархей, которых удалось вырастить в лаборатории — гетеротрофы, выделяющие молекулярный водород, предки эукариот, как выяснилось, были автотрофами и не выделяли водород, а потребляли его. Из этого следует, что те бактерии, с которыми вступили в симбиоз архейные предки эукариот, должны были водород выделять, а не потреблять, как предполагалось. Это согласуется с «водородной гипотезой» происхождения эукариот.
Появление эукариотической клетки — это и важнейшее событие в истории земной жизни, и одна из самых сложных головоломок для эволюционных биологов. Гипотетических сценариев происхождения эукариот предложено огромное количество, но не хватает данных для их проверки и детализации.
В последние годы в этой области наблюдается быстрый прогресс, связанный с достижениями сравнительной геномики и микробиологии, а главное, с открытием в 2015 году асгардархей — ближайших прокариотических родственников эукариот. «Элементы» внимательно следят за развитием этой темы и регулярно рассказывают о новых открытиях (см. ссылки в конце новости).
На сегодняшний день твердо установлено, что эукариотическая клетка произошла в результате симбиогенеза, то есть объединения нескольких прокариот в единый организм.
Центральной фигурой в этом объединении была архея из группы асгардархей. От нее эукариоты унаследовали большую часть компонентов систем репликации ДНК, транкрипции, синтеза белка и цитоскелета. Многие другие элементы эукариотической клетки, включая мембраны, мембранные белки и метаболические ферменты, унаследованы от бактерий, причем от разных. Часть — от альфапротеобактерии, ставшей предком митохондрий, часть — от какой-то другой или нескольких других бактерий.Поэтому, например, Евгений Кунин предполагает, что было как минимум два акта симбиогенеза. Сначала какая-то бактерия «проглотила» асгардархею. В получившейся химерной клетке асгардархея превратилась в клеточное ядро, а все мембраны стали бактериальными. Структура мембран бактерий и архей резко различается, а у эукариот все мембраны — бактериального типа, что неплохо объясняется предположением о «проглатывании» археи бактерией, а не наоборот. Затем эта химера «проглотила» альфапротеобактерию, которая дала начало митохондриям (см. Новое о происхождении эукариот: кто кого съел. Интервью с Евгением Куниным, «Троицкий вариант», 2024; Эукариотические клетки могли возникнуть в результате двойного эндосимбиоза, «Элементы», 10.07.2023).
Другие гипотетические сценарии учитывают, что прокариоты, в отличие от эукариот, — не большие мастера кого бы то ни было проглатывать. Зато у обоих представителей асгардархей, которых удалось культивировать в лаборатории и изучить под микроскопом, клетка образует длинные тонкие выросты (см. У асгардархей обнаружен сложный актиновый цитоскелет, «Элементы», 09.01.2023). Этими выростами можно плотно оплести симбионта, а потом выросты могут слиться. В результате симбионт станет внутренним, оставшиеся просветы между выростами образуют эндоплазматическую сеть, а центральная часть архейной клетки станет ядром (подробный разбор этой группы гипотез см. в новости Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019).
В той же новости обсуждается один смущающий факт, обозначенный как «проблема водорода». Выращенные в лаборатории асгардархеи оказались гетеротрофами, выделяющими молекулярный водород. Это плохо увязывалось с популярными гипотетическими сценариями эукариогенеза. Дело в том, что первым шагом на пути к симбиогенетическому слиянию археи с бактерией должна была быть синтрофия, то есть совместное проживание двух микроорганизмов, основанное на обмене необходимыми веществами. Наиболее правдоподобные и хорошо разработанные гипотезы о природе этой синтрофии предполагали, что архейный предок эукариот был автотрофом и нуждался в молекулярном водороде, который производился бактериальным партнером (обычно этого партнера трактуют как предка митохондрий, хотя на самом деле это могла быть и другая бактерия — та, что участвовала в первом из двух актов симбиогенеза).
Короче говоря, ожидалось, что асгардархеи потребляют H2, а оказалось, что они его, наоборот, выделяют. Исследователи, обнаружившие этот факт, справились с «проблемой водорода», предложив новый, довольно сложный сценарий поэтапного перехода от синтрофного сожительства гетеротрофной асгардархеи, выделяющей водород, с археями и бактериями, потребляющими водород, через симбиогенез к эукариотической клетке. То есть ученые, конечно, выкрутились, но осадочек остался (подробности см. в процитированной выше новости).
Новая статья китайских генетиков, опубликованная в журнале Nature, просто-напросто убирает «проблему водорода», позволяя вернуться к прежним идеям о том, что архейный предок эукариот был автотрофом, нуждающимся в молекулярном водороде.
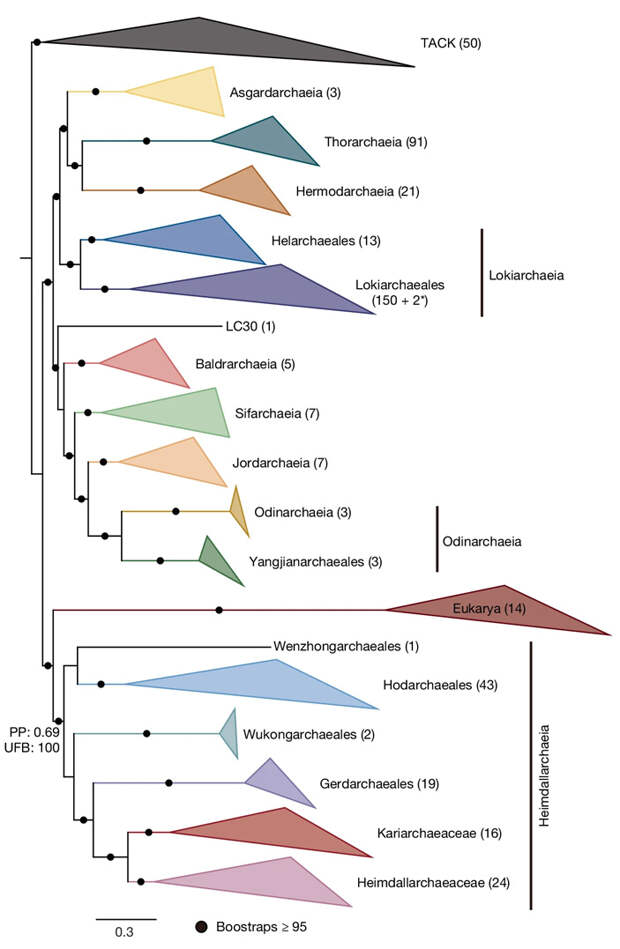
Главной целью исследования было уточнение эволюционного дерева асгардархей и места эукариот на нем. Как стало ясно уже в 2017 году, асгардархеи — это большая и разнообразная группа архей (см. Описан новый надтип архей, к которому относятся предки эукариот, «Элементы», 16.01.2017). Одной из ее веточек были архейные предки эукариот. Важно, что та подгруппа асгардархей, которая была впервые описана в 2015 году по ДНК из морских осадков возле гидротермальной системы «Замок Локи» в Северной Атлантике (см. Loki's Castle), и к которой относятся оба вида, которые удалось культивировать, — локиархеи — вовсе не самая близкая к эукариотам подгруппа. Самой близкой оказалась другая ветвь — хеймдалльархеи, известные пока только по ДНК, как и все остальные асгардархеи.
По мере того, как ученые секвенируют все новые и новые геномы асгардархей, их эволюционное дерево становится все более ветвистым, а место эукариот на нем постепенно уточняется. Однако до сих пор не было ясности в вопросе о том, являются ли эукариоты сестринской группой по отношению к хеймдалльархеям, или же архейные предки эукариот — это одна из веточек внутри кроны (см. Crown group) хеймдалльархей.
Этот вопрос важнее, чем может показаться на первый взгляд. Зная топологию дерева, можно реконструировать предковые состояния признаков в точках ветвления. Соответственно, можно понять, какой тип метаболизма был у первых представителей той ветви, которая привела к эукариотам. А от этого напрямую зависит, какой из гипотетических сценариев эукариогенеза более вероятен.
Результаты солидного исследования, опубликованного в Nature в 2023 году, свидетельствовали в пользу того, что архейные предки эукариот — одна из веточек кроны хеймдалльархей (L. Eme et al., 2023. Inference and reconstruction of the heimdallarchaeial ancestry of eukaryotes), сестринская по отношению к Hodarchaeales (это тоже подгруппа хеймдалльархей). Из этого, в свою очередь, следовало, что асгардархейные предки эукариот были гетеротрофами и в молекулярном водороде не нуждались, — как и оба представителя локиархей, культивированные в лаборатории.
Новая статья опровергает эти выводы. Китайские исследователи изучили ДНК из 40 образцов осадков мангровых болот и эстуариев из разных районов Китая. Полученные метагеномные данные позволили собрать 11 878 прокариотических геномов, из которых 223 принадлежат асгардархеям. Объединив их с геномами, опубликованными ранее, и убрав дубликаты, исследователи получили внушительную коллекцию, включающую геномы 411 видов (или штаммов) асгардархей. 136 из этих видов — новые. Описан также десяток новых родов и несколько семейств и отрядов.
Когда авторы начали разными методами строить по этим обширным геномным данным эволюционные деревья, они обнаружили, что одна недавно описанная группа, Njordarchaeales, ведет себя нестабильно. В зависимости от набора используемых для построения дерева генов и методических нюансов, Njordarchaeales то оказываются одной из веточек хеймдалльархей, то вообще выскакивают за пределы надтипа асгардархей и группируются с другим надтипом — Thermoproteati (альтернативное название — TACK). При ближайшем рассмотрении выяснилось, что геномы Njordarchaeales имеют химерную природу. Некоторые их фрагменты явно асгардархейные, но другие (и их даже больше) происходят от Thermoproteati. Не до конца понятно, то ли это действительно такая необычная химерная группа, возникшая в результате невероятно интенсивного обмена генами между двумя надтипами архей, то ли это результат ошибки при сборке геномов из кусочков ДНК, выделенных из проб. Так или иначе, для верности Njordarchaeales были исключены из дальнейшего анализа.
Следующей задачей было выяснение места эукариот (точнее, архейного предка эукариот) на обновленном дереве асгардархей. Деревья, построенные по генам, общим для асгардархей и эукариот, показали, что эукариоты уверенно занимают позицию, сестринскую по отношению ко всем хеймдалльархеям (рис. 1). Напомним, что в течение последних двух лет (после статьи L. Eme et al., 2023) считалось, что эукариоты находятся внутри кроны хеймдалльархей, а их сестринской группой являются Hodarchaeales — одна из подгрупп хеймдалльархей. По мнению авторов, ошибка была связана с химерной природой Njordarchaeales, которая искажала топологию эволюционного дерева, а также с меньшим числом учтенных геномов.
Уточненное дерево позволило при помощи тщательно откалиброванных молекулярных часов датировать основные события в эволюции асгардархей. Результаты показаны на рис. 2.

Получилось, что последний общий предок всех современных асгардархей жил 3,38 млрд лет назад (доверительный интервал: 3,72–3,06). Датировка последнего общего предка хеймдалльархей и локиархей — 3,16 млрд лет, а современные хеймдалльархеи начали дивергировать 2,72 млрд лет назад. Поскольку хеймдалльархеи — сестринская группа по отношению к эукариотам, получается, что обособление архейных предков эукариот должно было произойти раньше, чем 2,72 млрд лет назад (скорее всего, примерно 2,8 млрд лет назад). То есть задолго до кислородной революции (GEO на рис. 2), вероятно, в анаэробных условиях.
И, наконец, самое интересное: реконструкция предкового метаболизма на основе обновленного дерева. До сих пор считалось, что последний общий предок всех асгардархей, скорее всего, имел такой же тип метаболизма, как и два культивированных штамма локиархей. То есть был анаэробным гетеротрофом, потреблявшим аминокислоты и выделявшим в качестве отходов жизнедеятельности водород и короткоцепочечные жирные кислоты (ацетат, пропионат, бутират).
Однако новое исследование подтвердило альтернативную гипотезу, согласно которой предок асгардархей был хемоавтотрофом, зависимым от молекулярного водорода. У этого предка был полный набор ферментов древнего метаболического пути Вуда — Льюнгдаля (восстановительного пути ацетил-КоА; см. Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019). Кроме того, у него имелся набор железо-никелевых гидрогеназ. Это значит, что первые асгардархеи были автотрофами, фиксировали CO2 при помощи пути Вуда — Льюнгдаля, и нуждались для этого в молекулярном водороде.
В дальнейшем в некоторых линиях асгардархей, в том числе у локиархей и некоторых хеймдалльархей, часть ферментов пути Вуда — Льюнгдаля была утрачена — очевидно, вместе со способностью к автотрофному метаболизму. Оставшиеся ферменты этого пути обслуживают те или иные этапы гетеротрофного метаболизма. Например, у локиархей они служат для расщепления аминокислот, получаемых извне. Переход от автотрофного образа жизни к гетеротрофному, по-видимому, неоднократно происходил у разных асгардархей.
Но не у предков эукариот. У последнего общего предка хеймдалльархей, как и у последнего общего предка хеймдалльархей и эукариот, сохранился полный набор ферментов пути Вуда — Льюнгдаля. Авторы статьи L. Eme et al. (2023) этого не увидели, потому что в их распоряжении было гораздо меньше геномов хеймдалльархей, причем как раз в тех геномах, которые они анализировали, часть ферментов этого пути отсутствует. Однако во многих новых геномах хеймдалльархей, впервые изученных в обсуждаемой работе, есть полный набор этих ферментов, а также гидрогеназы. Кроме того, у первых хеймдалльархей, как и у первых представителей эукариотной линии, были ферменты фосфотрансацетилаза и ацетат-киназа. Это значит, что они были ацетогенами: производили ацетат в качестве побочного продукта своего автотрофного метаболизма.
Таким образом, архейный предок эукариот был асгардархеей, родственной хеймдалльархеям, но не был представителем кроновых хеймдалльархей. Он отделился базальнее кроны этой группы. Он был строгим анаэробом, потому что некоторые его ферменты крайне чувствительны к кислороду и не смогли бы работать даже в присутствии небольших концентраций этого яда. Он был хемоавтотрофом, фиксировал CO2 при помощи пути Вуда — Льюнгдаля, использовал H2 в качестве восстановителя и выделял ацетат как конечный продукт метаболизма.
Эти выводы позволяют сузить круг гипотетических сценариев эукариогенеза. Главный вопрос — откуда архейный предок эукариот получал необходимый ему водород? Поначалу он мог не заморачиваться с синтрофией и использовать H2, образующийся в результате серпентинизации — геологического процесса, который и сегодня обеспечивает водородом некоторых нуждающихся в нем микробов, живущих в гидротермальных условиях (см. Serpentinization). На молодой Земле серпентинизация шла очень активно, однако к концу архейского эона (завершившегося 2,5 млрд лет назад) ее темпы резко упали (см. J. Leong et al., 2021. Decreasing extents of Archean serpentinization contributed to the rise of an oxidized atmosphere). Теперь уже имело смысл подумать о синтрофном партнере.
Согласно известной «водородной гипотезе» происхождения эукариот (W. Martin, M. Müller, 1998. The hydrogen hypothesis for the first eukaryote), поставщиком водорода для архейного предка была альфапротеобактерия — предок митохондрий. Действительно, некоторые альфапротеобактерии (например, пурпурные несерные альфапротеобактерии, практикующие аноксигенный фотосинтез) выделяют H2.
Авторы подчеркивают, что их результаты хорошо согласуются с «водородной гипотезой» Мартина и Мюллера.
Впрочем, не будем забывать, что в симбиогенезе участвовало более одного вида бактерий. Реальная история синтрофных отношений архейного предка эукариот с различными бактериями наверняка была более сложной, чем однократное объединение с альфапротеобактериальным предком митохондрий.
В этих взаимоотношениях могли быть задействованы сульфатредуцирующие бактерии. С ними, как известно, вступают в синтрофные отношения локиархеи (см. Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019). Некоторые сульфатредукторы утилизируют ацетат, от которого архейному предку эукариот нужно было избавляться. Они утилизируют также сульфат, производимый пурпурными несерными альфапротеобактериями. На возможное участие в эукариогенезе сульфатредуцирующих бактерий (или их родичей из группы дельтапротеобактерий) намекают и геномные данные. Однако сульфатредукторы не производят водород, а, наоборот, потребляют его. Значит, нашему воображаемому синтрофному комплексу не помешали бы дополнительные поставщики водорода. На эту роль могли бы подойти гетеротрофные бактерии-бродильщики (см. Fermentation), сбраживающие сахара и выделяющие при этом водород. Не исключено, что эукариоты унаследовали от таких синтрофных партнеров кое-что полезное, например, ферменты гликолиза — основного энергетического процесса эукариотической цитоплазмы.
Таким образом, обсуждаемая работа представляет собой важный шаг на пути к сужению круга вероятных сценариев эукариогенеза. Будем надеяться, что дальнейшее накопление данных позволит уже в ближайшем будущем прийти к какому-то одному хорошо обоснованному и детализированному сценарию.
Источник: Jiawei Zhang, Xiaoyuan Feng, Meng Li, Yang Liu, Min Liu, Li-Jun Hou & Hong-Po Dong. Deep origin of eukaryotes outside Heimdallarchaeia within Asgardarchaeota // Nature. 2025. DOI: 10.1038/s41586-025-08955-7.
См. также:
1) Новое о происхождении эукариот: кто кого съел. Интервью с Евгением Куниным // «Троицкий вариант», 2024.
2) Эукариотические клетки могли возникнуть в результате двойного эндосимбиоза, «Элементы», 10.07.2023.
3) У асгардархей обнаружен сложный актиновый цитоскелет, «Элементы», 09.01.2023.
4) Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019.
5) Описан новый надтип архей, к которому относятся предки эукариот, «Элементы», 16.01.2017.
6) Полиплоидность предков эукариот — ключ к пониманию происхождения митоза и мейоза, «Элементы», 14.06.2016.
7) Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016.
8) Новооткрытый микроб заполняет брешь между прокариотами и эукариотами, «Элементы», 12.05.2015.
9) А. В. Марков, А. М. Куликов, 2009. Происхождение эукариот как результат интергационных процессов в микробном сообществе.
Свежие комментарии