
Птичье пение заметно отличается не только у разных видов, но нередко и у разных популяций одного вида. Такие особенности называют песенными диалектами: они формируются подобно диалектам в человеческих языках в процессе культурной эволюции. Как и в случае с речью, полноценное освоение песни требует обучения, но эта способность имеет и врожденный компонент — в частности, склонность к запоминанию песни своего вида. Молодую птицу окружает множество звуков, а значит, во время обучения ей нужно как-то опознавать «своих». Это может быть особенно актуально там, где соседствуют два близких вида с похожей вокализацией — такие как мухоловка-пеструшка (Ficedula hypoleuca) и мухоловка-белошейка (Ficedula albicollis). Авторы нового исследования решили проверить, связано ли предпочтение песни своего вида с присутствием близкого «родича» и отбором против гибридов. Оказалось, что птенцы мухоловок-пеструшек, еще не обучившиеся песне, одинаково успешно опознают пение своего вида как в зоне совместного обитания с белошейкой, так и там, где ее нет. Более того, незадолго до вылета из гнезда птенцы пеструшек отдают явное предпочтение диалекту той популяции, где они появились на свет, а реакция на диалекты других популяций по интенсивности не превосходит реакцию на пение мухоловки-белошейки. Есть веские основания полагать, что такие предпочтения не связаны с обучением: по-видимому, не только сама песня, но и ее локальные особенности из культурной традиции успели превратиться в генетически наследуемый признак.
Певчие воробьиные птицы (Passeri) — одна из немногих групп животных, которые способны выучивать новые звуки и использовать их в своей коммуникации.
Большинство вокальных сигналов, будь то лягушачье кваканье или кошачье мяуканье — врожденные: животные издают их, даже если никогда в жизни не слышали своих сородичей. Из млекопитающих способность к вокальному обучению известна у грызунов, хоботных, китообразных, ластоногих и некоторых приматов, включая, разумеется, и людей. У птиц, кроме певчих воробьиных, обучаться звукам могут колибри и попугаи (см. Vocal learning).Звуковые сигналы, которые осваиваются в процессе обучения, могут меняться очень быстро — на их развитие влияют процессы культурной эволюции. Ошибки копирования или новации, вводимые отдельными особями, приводят к появлению новых вариантов сигналов. В разных популяциях ход этих изменений скорее всего будет неодинаковым, а значит, при условии даже частичной изоляции различия в их вокализации будут накапливаться — вплоть до того, что носители разных диалектов перестают «понимать друг друга». Такой процесс может существенно ускорить разделение ранее единого вида на несколько сестринских.
Но чтобы общаться с себе подобными, нужно разговаривать «на их языке». Поэтому животным с вокальным обучением нужны специальные механизмы, которые обеспечивали бы узнавание и запоминание звуков своего вида, а не какого-то другого. Конечно, бывает и так, что птицы воспроизводят «чужие» звуки: виды-имитаторы вставляют в свои песни заимствованные элементы (см. картинку дня Птичье кваканье), а в условиях эксперимента воспроизводить чужую песню могут и те виды, которым заимствования не свойственны. Но это возможно только в случае, если молодых птиц изолируют от звуков сородичей; и даже тогда песня может иметь заметный «акцент» (см. В птичьей песенке наследуются интервалы, а сами звуки выучиваются, «Элементы», 13.12.2016). Если же птицы слышат пение своего вида, они будут обучаться именно ему, игнорируя прочих «певцов». Значит, вокальное обучение накладывается на некую наследственную «матрицу», а способность выделять песни своего вида в окружающем звуковом фоне определяет врожденный «фильтр».
Впрочем, в случае близких видов межвидовые барьеры не столь уж непроницаемы. Например, восточный соловей (Luscinia luscinia) в зонах симпатрии (совместного обитания двух видов) перенимает песни у южного соловья (Luscinia megarynchos; см. И. М. Марова и др., 2015. Сравнительный анализ дифференциации песни восточного (Luscinia luscinia) и южного (Luscinia megarhynchos) соловьев на пространстве их ареалов). Песни этих видов довольно близки по своему звучанию, так что такие заимствования можно списать на несовершенство врожденных «фильтров», хотя не исключено, что имитация имеет и функциональный смысл: с помощью нее самцы могут расширять свой вокальный репертуар или обеспечивать охрану территории от другого вида. Впрочем, возможны и негативные последствия: исполнение песни другого вида повышает риск гибридизации, а гибридное потомство нередко оказывается бесплодным.
Итак, способность к безошибочному узнаванию песни своего вида особенно актуальна в зонах симпатрии с близким родственником. Поэтому можно предположить, что именно там она будет развита наилучшим образом. Авторы нового исследования проверили это предположение на мухоловке-пеструшке (Ficedula hypoleuca) — пожалуй, самом популярном виде Старого Света для всевозможных поведенческих и экологических исследований. Мухоловки-пеструшки довольно многочисленны в лесах Европы, и они охотно занимают искусственные гнездовья — дуплянки. Это сильно облегчает поиск гнезд, мечение и индивидуальное слежение за птицами. В Центральной и Восточной Европе ареал пеструшки пересекается с ареалом мухоловки-белошейки (Ficedula albicollis): внешне эти виды очень похожи (рис. 1), но их потомство бесплодно, поэтому птицам важно избегать «смешанных браков».
Песня мухоловки-пеструшки состоит из фраз длительностью 2–2,5 с. Каждая из них обычно состоит из 7–10 коротких повторяющихся единиц, которые называют слогами (в других работах — элементами или фигурами): каждый слог включает один-два, реже несколько звуков, разделенных короткими паузами (до 30 мс). Высокие и низкие слоги часто чередуются, создавая узнаваемое звучание песни: его изображают как «клю-чик, клю-чик, вер-ти, кру-ти, кру-ти» (жирным шрифтом выделены высокие слоги; пример песни мухоловки-пеструшки). Песня белошейки очень близка к песне пеструшки по частотным характеристикам, но состоит из более продолжительных свистовых элементов и в целом длиннее. Звучит она примерно как «фи-циии-цзе-триии-трю-трюю»; резкие перепады высоты следующих друг за другом нот в ней редки (пример песни мухоловки-белошейки).
Молодые птицы обоих видов чаще всего занимают гнездовые территории в 5–20 км от того места, где они появились на свет. Поэтому, хотя между популяциями и нет непреодолимых географических барьеров, в разных регионах формируются специфичные песенные диалекты. Как и большинство певчих воробьиных птиц, мухоловки-пеструшки начинают обучаться песне в возрасте приблизительно 20 дней. Во всяком случае, все то, что молодые птицы слышали в более раннем возрасте, никак не влияет на их будущий вокальный репертуар. При этом реакция на песню своего вида проявляется намного раньше: слуховая система птенцов начинает воспринимать звуки песни на 4–5 день жизни, а способность отличать пение своего вида формируется на 9–11 день. Эксперименты провели с птенцами в возрасте 11 или 12 дней; позже, в 13–18 дней, они уже покидают гнездо. Это намного меньше того возраста, в котором мухоловки начинают запоминать элементы своей будущей песни, поэтому исследователи предполагали, что реакция на стимулы будет сугубо врожденной, не связанной с предшествующим опытом (это предположение проверили в ходе специальных тестов, о которых будет сказано ниже).
В ходе эксперимента сравнивали интенсивность реакции птенцов на песни, типичные для их родной популяции, и на иные варианты песен — характерные для других популяций вида или мухоловки-белошейки. Суммарно в экспериментах поучаствовали 1722 птенца мухоловок-пеструшек из 372 выводков и 7 популяций. Почти все они были аллопатричными, и только популяция шведского острова Эланд находилась в зоне симпатрии с белошейкой (рис. 2, A).

Для экспериментов отобрали 245 записей от 168 особей двух видов мухоловок в районах исследований. Чтобы выявить вокальные особенности разных популяций, авторы применили специальный алгоритм в программе Luscinia, оценивающей сходство песен: при этом учитываются как частотные характеристики, так и ритм исполнения. Результатом стала диаграмма рассеяния, отражающая степень сходства песен разных самцов (рис. 2). Как видно из рисунка, диалекты не имеют четких границ, поэтому за тот или иной диалект принимали область, в которую попадали 95% песен соответствующей популяции. А вот песни пеструшек и белошеек по своим параметрам расходятся довольно четко — хотя некоторые из «пеструшечьих» песен в симпатрической популяции оказались ближе к песням белошейки. Авторы не говорят о причинах такого сходства, но можно предположить, что оно связано с копированием песен близкого вида в зоне симпатрии. Интересно, что такое копирование характерно только для мухоловки-пеструшки, даже в тех регионах, где она значительно превосходит по численности белошейку (см. А. П. Вабищевич, Н. А. Формозов, 2008. Изменчивость песни мухоловки-пеструшки (Ficedula hypoleuca) в зоне симпатрии с мухоловкой-белошейкой (Ficedula albicollis)). Любопытно, что пеструшки воспроизводят звуки чужой песни в своем собственном «пеструшечьем» ритме — похожую закономерность обнаружили в экспериментах на разных видах амадин, которые обучались песне другого вида (см. В птичьей песенке наследуются интервалы, а сами звуки выучиваются, «Элементы», 13.12.2016).
В ходе эксперимента подопытным птенцам, которых временно изымали из гнезд, в случайном порядке проигрывали три стимула длительностью в одну минуту — песню диалекта их собственной популяции, песню другого диалекта и песню мухоловки-белошейки. В экспериментальных укрытиях, имитирующих гнездо, были установлены видеокамеры и микрофоны, которые фиксировали реакцию птенцов на проигрывание песен. Каждое проигрывание предварялось минутой тишины, так что весь эксперимент занимал 6 минут. Анализируя записи, один из авторов работы подсчитывал различные проявления активности птенцов: пищевые позывки — с их помощью птенцы выпрашивают корм у родителей, задирание головы вверх, раскрывание клюва и подпрыгивания. Наиболее используемыми и удобными для подсчета оказались пищевые позывки — так что в дальнейшем анализе в качестве показателя интенсивности реакции использовали число позывок во время проигрывания песни. Поскольку реакции птенцов из одного гнезда были сходными, результаты усреднили для каждого выводка.
Оказалось, что во всех популяциях птенцы реагируют и на «родную» песню, и на песню близкого вида. Этот результат вполне ожидаем, если учесть, что по частотным характеристикам те и другие практически идентичны. Однако интенсивность реакции на родную песню была достоверно выше, чем на песню белошейки (рис. 3). При этом в симпатричной популяции эта разница была не больше, чем в аллопатричных. Не зависела она и от расстояния до зоны симпатрии. А значит, успех распознавания видоспецифичной песни не связан с «эволюционным опытом» контакта с близким видом.
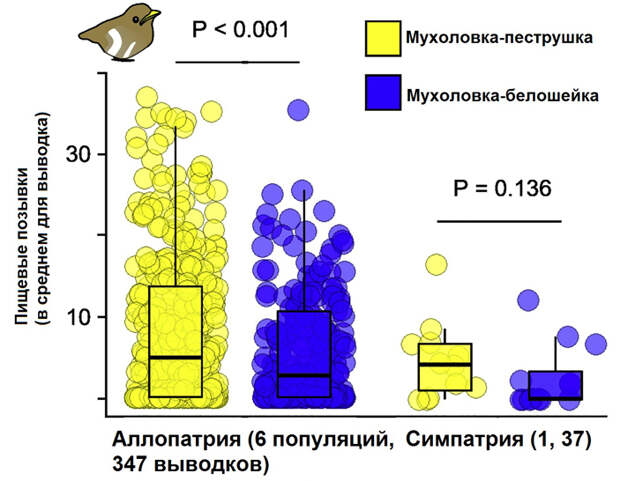
Конечно, мастерство распознавания могло оттачиваться и под влиянием взаимодействия с каким-нибудь другим повсеместно распространенным видом, например большой синицей (Parus major). Но это не объясняет самый неожиданный результат (рис. 4): в возрасте 12 дней птенцы отдавали явное предпочтение песне «родного» диалекта, тогда как песни из других популяций — не важно, соседних или удаленных — по интенсивности реакции не превосходили песни другого вида! Все это означает, что врожденный «фильтр» птенцов настроен именно на локальный диалект, и различение песни других видов — не причина, а полезный сопутствующий эффект такой настройки. Конечно, приобретенный опыт тоже играет свою роль в межвидовом узнавании, но он приходит уже в более зрелом возрасте: здесь авторы ссылаются на работы, которые демонстрируют, что взрослые мухоловки из симпатрических популяций — как самцы, так и самки — отличают близкий вид лучше, чем птицы из аллопатричных популяций (M. N. Verzijden et al., 2012. The impact of learning on sexual selection and speciation; L. Bliard et al., 2021. The role of introductory alarm calls for song discrimination in Ficedula flycatchers).
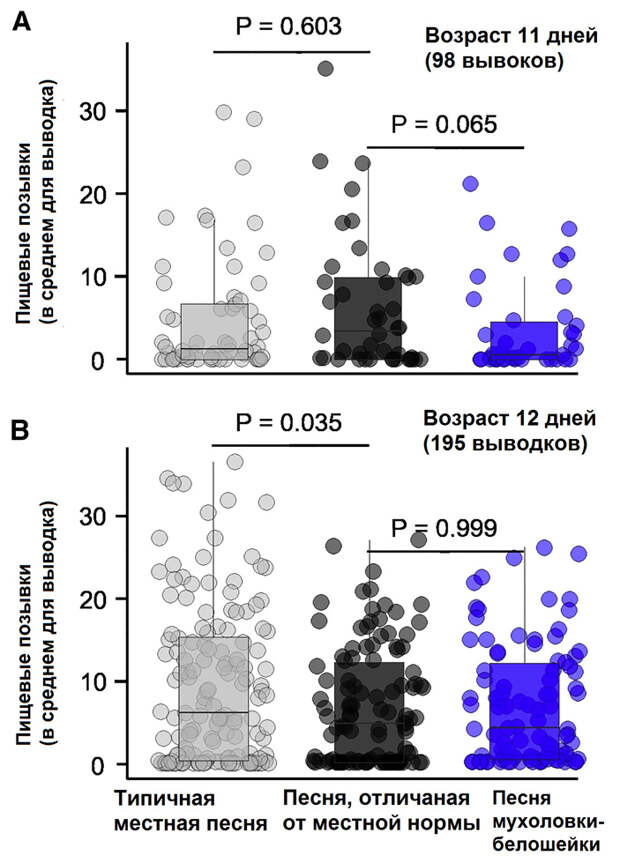
Конечно, полностью исключить возможность обучения у птенцов все же нельзя: ведь они росли в естественной обстановке, где вполне могли слышать пение своего отца или других самцов. Однако авторы отмечают, что, во-первых, о запоминании песни в таком раннем возрасте у этого вида ничего не известно, а во-вторых, во время выкармливания большинство самцов поет редко. Чтобы проверить, как акустический опыт птенцов влияет на узнавание песни, в одной из популяций (окрестности зоологической станции Товеторп в Швеции; TO на рис. 2) авторы записывали звуковое окружение 28 гнездовых дуплянок. Записи делали в утреннее время (до 8 часов), когда мухоловки поют наиболее активно; у каждой дуплянки звуковой фон записывали от 4,4 до 22,3 часов (медиана 12,5 часов).
По сонограммам этих записей подсчитали число песен мухоловок-пеструшек, тем самым оценив, как часто птенцы в каждом гнезде слышали пение своего вида. Можно представить, насколько трудоемко было бы обрабатывать эти записи вручную, учитывая, что их суммарная продолжительность — больше двух недель чистого времени. К счастью, сейчас уже появились программы для автоматического выделения звуков тех или иных животных; одной из таких программ и воспользовались авторы статьи. Разумеется, все обнаруженные песни проверили вручную для контроля возможных ошибок (точность обнаружения составила примерно 90%). Затем обнаруженные песни сравнили с популяционной нормой по той же методике, что применялась для выявления диалектной структуры — это позволило оценить, насколько типичные для данной популяции песни слышал выводок от своего отца (или других местных птиц — хотя у такого территориального вида, как мухоловка-пеструшка, они вряд ли часто поют у чужого гнезда).
Если бы повышенная реакция птенцов на песни местного диалекта была связана с обучением, то большую «разборчивость» должны проявлять те птенцы, которые слышали пение чаще. Соответственно, чем больше слышанные ими песни похожи на популяционную «норму», тем сильнее их предпочтение местного диалекта. Однако результаты оказались едва ли не противоположными: у птенцов, чаще слышавших пение, предпочтение локального диалекта было выражено даже меньше, чем у тех, кто слышал его реже. Более того, птенцы слабее реагировали на пение, похожее на то, которое они слышали в гнезде, а предпочтение локального диалекта никак не зависело от того, насколько пение их отца близко к популяционной норме. Авторы отмечают, что в период ухода за потомством наиболее активно поют те самцы, которые стремятся привлечь еще одну самку: среди мухоловок-пеструшек обычна би- и даже тригамия, а также внебрачные спаривания. По-видимому, у птенцов, которым часто приходилось слышать пение рядом с гнездом, попросту развивалось привыкание, поскольку его звуки переставали ассоциироваться с появлением родителя с кормом.
Все это позволяет заключить, что хотя птенцы мухоловок и могут слышать «родной» диалект, их предпочтения вряд ли определяются таким опытом. Впрочем, авторы оговариваются, что для окончательного подтверждения врожденного характера диалектных «пристрастий» потребуются экспериментальные исследования с птенцами, изъятыми из гнезд сразу после вылупления и выращенными в звуковой изоляции.
Происхождение поведенческих моделей — проблема, волновавшая ученых с самого начала развития эволюционных теорий. Трудно представить, что сложное адаптивное поведение формируется в результате череды случайных мутаций. А если каким-то животным и удается его освоить, передать этот опыт потомкам они могут лишь путем обучения, а не генетически. Идея, позволяющая связать обучение и эволюцию, была сформулирована еще в конце XIX века — сегодня эта концепция известна как эффект Болдуина. Суть его такова: если у животного есть способность к освоению того или иного навыка, повышающего приспособленность, то преимущество получат те особи, которые обучаются данному навыку более эффективно. Например, они могут — благодаря той или иной мутации — обзавестись врожденными «задатками», способствующими запоминанию песни своего вида, а не какой-то другой. Потомки успешной особи унаследуют полезную мутацию, которая помогает осваивать культурно передаваемый признак. Таким образом, поведение животного задает направление естественного отбора и делает его значительно более эффективным, а признак, который изначально приобретался в ходе обучения, может со временем закрепиться и в генетической наследственности. По результату эффект Болдуина напоминает «ламарковское» наследование приобретенных признаков, но реализуется он за счет «дарвиновского» механизма.
Хоть авторы обсуждаемой статьи и не касаются этих вопросов, они подчеркивают, что их исследование демонстрирует сам факт «превращения» культурно наследуемой черты в наследуемый признак. К слову, одна из гипотез, которой объясняют феномен песенных диалектов — это гипотеза локальной адаптации: особи, происходящие из местной популяции, адаптированы к местным условиям лучше «чужаков», а значит, самкам, выбирающим пару, следует предпочесть именно их (см. В. В. Иваницкий, 1989. Коммуникация у животных: теории и факты). Это может объяснять эволюционную выгоду диалектных предпочтений, хотя как они влияют на будущий выбор партнера, еще предстоит разобраться. В эволюционной перспективе, такие предпочтения могут ускорить дивергенцию популяций и разделение ранее единого вида на несколько. А значит, вокальное обучение может быть одной из причин поразительного видового богатства певчих воробьиных птиц.
Источник: David Wheatcroft, Louis Bliard, Myriam El Harouchi, David López-Idiáquez, Tiia Kärkkäinen, Fanny-Linn H. Kraft, Jaime Muriel, Samyuktha Rajan, Tomas Tuvillo, Malcolm D. Burgess, Alejandro Cantarero, Toni Laaksonen, Jesús Martínez-Padilla, Marcel E. Visser, Anna Qvarnström. Species-specific song responses emerge as a by-product of tuning to the local dialect // Current Biology. 2022. DOI: 10.1016/j.cub.2022.09.063.
Антон Морковин
Свежие комментарии