
Уже на ранних стадиях своей эволюции цветковые растения научились использовать насекомых для переноса пыльцы. Вскоре это в буквальном смысле привело к их процветанию и развитию тесного мутуалистического симбиоза энтомофильных цветковых с шестиногими опылителями. Сейчас глобальные экологические проблемы угрожают этой многомиллионнолетней связи — из-за сокращения численности насекомых-опылителей, в том числе пчел. Чтобы оценить масштабы сопутствующих эволюционных изменений у растений, авторы статьи в журнале New Phytologist «воскресили» полевые фиалки (Viola arvensis), семена которых были собраны 20–30 лет назад, и сравнили их с современными фиалками из тех же экосистем неподалеку от Парижа. Оказалось, что во всех четырех изученных локациях на фоне недостатка опылителей у V. arvensis произошли характерные изменения генеративных органов — усилился так называемый «синдром самоопыления». Размер привлекающих насекомых частей цветков уменьшился и они стали вырабатывать меньше нектара. В результате их более невзрачные цветки реже обращают на себя внимание насекомых — это подтвердили эксперименты со шмелями, которые предпочитали «воскрешенные" фиалки современным. Более того, фактическая частота самоопыления у V. arvensis резко возросла — в среднем на 27%.
Цветковые или покрытосеменные растения (Angiospermae) намного моложе других групп высших растений. Предполагают, что активное образование таксонов цветковых началось около 100 миллионов лет назад — именно тогда стартовало их триумфальное распространение по Земле с освоением все новых территорий и разнообразных экологических ниш. Важным «катализатором» эволюции цветковых уже на ранних этапах стал тесный мутуалистический симбиоз с насекомыми-опылителями, который обеспечило появление энтомофильных цветков. Союз цветка и опылителя привел к развитию ряда взаимных адаптаций с обеих сторон, в том числе к переходу некоторых насекомых (в частности, пчел (Anthophila)) на питание пыльцой и нектаром — специализированным «угощением», предоставляемым цветами.
В результате подавляющее большинство (около 80%) современных цветковых растений энтомофильны. Остальные используют в качестве вектора для переноса пыльцы других животных, ветер (анемофилия) или даже воду (гидрофилия). Заметная часть покрытосеменных способна к самоопылению (автогамии) с последующим оплодотворением. Считается, что самоопыление всегда возникает вторично, в результате перехода от перекрестного опыления в условиях, где оно затруднено: скажем, в экстремальных северных экосистемах, пустынях, горах и т. д. Таким образом, опыление собственной пыльцой — это «запасной» способ размножения, который страхует растения на случай когда перекрестное опыление невозможно. В то же время самоопыление (и вообще самооплодотворение организмов) в долговременной перспективе может быть чревато рядом генетических и эволюционных последствий.
Еще Чарлз Дарвин писал, что «природа питает отвращение к постоянному самооплодотворению», не забыв добавить следом, что без слова «постоянное» фраза теряет смысл (Ч. Дарвин, Действие перекрестного опыления и самоопыления в растительном мире; в издании 1939 года приведенная цитата на стр. 17). Поэтому в природе самоопыление обычно сочетается с «нормальным» перекрестным опылением — например, с помощью насекомых.Однако глобальные экологические проблемы, такие как деградация экосистем, сокращение биоразнообразия и загрязнение окружающей среды, привели к тому, что опылителей становится все меньше (см., например, Проблема жъжъь. Что такое «кризис опыления» и как с ним быть). Тревожная тенденция прослеживается не менее полувека и связана в том числе с активным использованием пестицидов. Ситуация угрожает устоявшейся экологической связи между цветковыми и насекомыми-опылителями. Более того, под вопросом оказываются экосистемные услуги, которые те оказывают посевам, что грозит негативными последствиями для сельского хозяйства.
Недостаток попадающей в цветок извне пыльцы считают одной из главных причин перехода растений к самоопылению. Смена стратегии опыления не раз описана в дикой природе, хотя речь как правило идет о крупных временных масштабах. Однако в условиях антропогенной нагрузки на экосистемы (прежде всего дефицита опылителей) насекомоопыляемые растения могут переключиться на самоопыление гораздо быстрее — за считанные десятилетия. Отдельные подтверждения тому имелись и раньше, а теперь феномен на примере полевой фиалки (Viola arvensis) подробно описали в своей недавней статье в журнале New Phytologist ученые из Университета Монпелье (Франция).
Исследователи использовали остроумный подход — «экологическое воскрешение» (resurrection ecology), предполагающий оживление (в данном случае — проращивание) семян, спор, яиц и так далее, которые долгое время провели в состоянии покоя. В отличие от длительных непрерывных экспериментов, такая методология позволяет разделить влияние генетических и экологических факторов на фенотип. Объектом исследования стала полевая фиалка Viola arvensis (рис. 1) — небольшое однолетнее или двухлетнее растение с обширным ареалом, который охватывает Восточную Европу и значительную часть Сибири. В Северной Америке V. arvensis стала инвазивным видом (A. Scoppola et al., 2022. Classical Morphometrics in V. arvensis and V. kitaibeliana (Viola sect. Melanium) Reveals Intraspecific Variation with Implications for Species Delimitation: Inferences from a Case Study in Central Italy). По-видимому, исходно эта фиалка росла на лугах и вдоль рек, однако сейчас оказалась также широко распространенным сорняком, вредителем многих посевов.
Генеративная сфера полевой фиалки представлена зигоморфным цветком размером до 1 сантиметра и более. Венчик состоит из 5 лепестков неправильной формы, причем губа (лат. labium) — крупный и непарный нижний лепесток — приспособлена для «посадки» насекомого и даже имеет характерные темные черточки (nectar guide), которые направляют посетителя цветка в нужную сторону, то есть к нектару. Фиалки также имеют шпорец — полый вырост, предназначенный для хранения нектара. Все эти признаки очевидно делают цветок привлекательным для насекомого и указывают на его энтомофильность. В то же время V. arvensis и фиалки в целом приспособлены к самоопылению, приводящему к самооплодотворению и развитию семян и новых растений, которые имели единственного родителя.
Исследованные фиалки росли в диком виде на севере Франции, сравнительно недалеко от Парижа, в следующих локациях: Круи (Crouy), Люи (Lhuys), Коммени (Commeny) и Герн (Guernes). Местные экосистемы относятся к числу сильно пострадавших от сокращения числа насекомых-опылителей. Семена V. arvensis в Круи и Люи были собраны в 1990-х, в Коммени и Герн — в 2000-х годах. Исходную выборку семян получали случайно, используя для этого не менее 100 материнских растений. В феврале 2021 года авторы нового исследования посетили те же экосистемы — максимально близко к месту первых сборов — и вновь получили семена V. arvensis, уже современных.
Чтобы «воскресить» фиалки, которые росли 20 или даже 30 лет назад, и корректно сравнить их с современными растениями, ученые для начала вырастили «освежающее поколение» F0 (refresher generation). Хранившиеся семена прорастили — их всхожесть составила 50–65%. Аналогичные растения поколения F0 получили и для современных фиалок, которые были собраны в дикой природе в виде проростков. Данное поколение не использовали в экспериментах, чтобы избежать искажений, связанных с хранением семян и индивидуальными особенностями материнских растений. Все фиалки выращивали рядом в непроницаемых для насекомых клетках, а затем подвергли контролируемому опылению. Среди растений поколения F0 случайным образом выбрали по 20 родительских растений (для каждой локации, в общей сложности 80), используя которые получили по 20 родственных групп («семей») поколения F1. Для этого внутрь клеток помещали ульи земляных шмелей (Bombus terrestris) — сейчас такие может приобрести любой желающий. В результате получили семена «экспериментального» поколения F1 (из каждой «семьи» выбирали по 30 семян), из которых вырастили растения, использованные для морфометрии и опытов с участием опылителей — в их роли выступили те же шмели. Наконец, из каждой «семьи» вновь было отобрано по пять проростков (или меньше в случае низкой всхожести) — такие сестринские растения использовали для получения нового набора «семей». Поскольку V. arvensis склонна к самоопылению, большая часть фиалок были инбредны — авторы подтвердили это с помощью микросателлитных маркеров. В общей сложности в экспериментах были задействованы 792 растения.
Для начала биологи охарактеризовали популяционную генетику растений поколения F0 (как «более близкого» к природной популяции) на основе 11 микросателлитных маркеров. Так они рассчитали относительное число аллелей (англ. allelic richness), ожидаемый уровень гетерозиготности (expected heterozygosity) и ее снижение (heterozygosity deficiency), а также выяснили, как изменилось генетическое разнообразие популяций V. arvensis со временем. Исходя из снижения (недостаточности) гетерозиготности также получены оценки частоты самоопыления.
Далее работали с «экспериментальным» поколением F1. Были описаны морфологические и некоторые другие признаки как «воскрешенных», так и современных экземпляров V. arvensis, что позволило выявить их фенотипические различия. Генеративные органы охарактеризовали следующими признаками (рис. 2): длина венчика, ширина губы, примерная оценка площади цветка (как произведение этих двух чисел), длина шпорца, длина чашечки, отношение длины чашечки к длине венчика (в миллиметрах), число «направляющих черточек» (nectar guides) на губе, длительность цветения (в днях), дата зацветания, среднее число одновременно открытых цветков на растении (floral display) и объем нектара (в микролитрах). Авторы отметили, что с целью описания морфологии цветков V. arvensis оптимально рассматривать те 5 цветков на самом крупном стебле, которые открываются первыми. В общей сложности они описали около 4000 цветков.
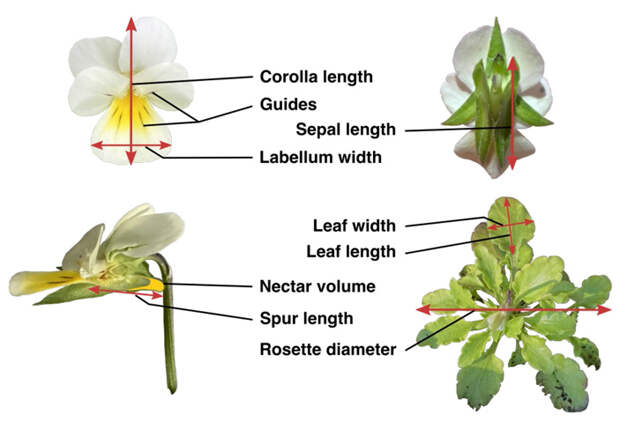
Также авторы охарактеризовали вегетативную сферу растений: диаметр прикорневой розетки, длину и ширину самого крупного листа (в миллиметрах), оценили его площадь (как произведение этих двух чисел), а также общую сухую биомассу после отмирания. Затем они выяснили, как происходит образование плодов у V. arvensis в присутствии насекомых-опылителей и без них. Для этого они пометили самые молодые и способные к опылению цветки и те, которые были заведомо слишком стары для этого. Таким образом, полученные в результате переноса пыльцы насекомым плоды формировались позже и были отличимы. Как и прежде, растения помещали в клетки, через стенки которых не могут пролезть насекомые, внутрь которых ставили шмелиные ульи. Опылителям давали по меньшей мере 2 часа на посещение цветков, после чего фиалки убирали. Таким образом авторы узнали соотношение семян, полученных за счет самоопыления и при помощи насекомых, а также их вес.
Далее ученые выяснили, насколько привлекательны для насекомых цветки из разных "времен» и локаций. Шмелиные ульи и фиалки вновь поместили в «непроницаемые» для насекомых клетки: по 20 экземпляров родом из одной и той же локации, среди которых 10 «воскрешенных» (из предковой популяции) и 10 современных. В ходе эксперимента наблюдатель выбирал одного из занятых опылением шмелей и замечал, какие цветки он посещает за 10–15 минут своей работы. В среднем за это время шмель посещал 15,72 растения — за время всех экспериментов «обойденными» оказались 124 фиалки (всего 580, считая 6–8 повторностей). Привлекательность растения для шмелей оценили, поделив число посещений его цветков к общему числу посещений для этого насекомого за один эксперимент.
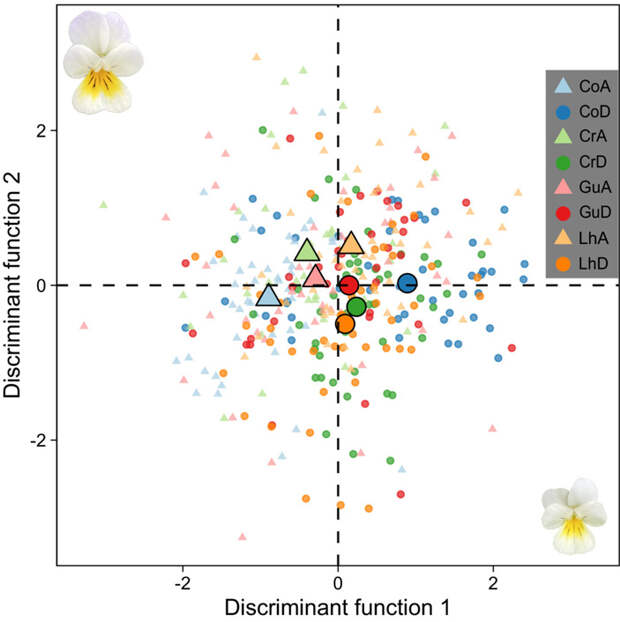
Собранные данные подвергли статистической обработке. С помощью линейного дискриминантного анализа (linear discriminant analysis, LDA) авторы описали различия 8 экспериментальных групп фиалок по 9 морфологическим признакам (рис. 3). Задача LDA — найти линейную комбинацию признаков, которая позволяет выделить и описать группы в анализируемых данных. Изменения отдельных признаков провели с помощью смешанных линейных моделей (univariate linear mixed models LMM). Каждый из признаков считали зависимой переменной (response variable), тогда как принадлежность к «предковой» или «потомковой» группе, одной из 4 локаций, сочетание того и другого, а также отнесение к выведенным авторами «семьям» являлись независимыми категориальными переменными. Если авторы замечали существенный эффект возраста (то есть принадлежности к «предковой» или «потомковой» популяциям), они дополнительно проверяли его с помощью t-теста с поправкой Бонферрони.
Наконец, результаты экспериментов с насекомыми анализировали с помощью обобщенных смешанных линейных моделей (GLMM). Из рассмотрения исключили 124 фиалки, которые не посетило ни одно насекомое. Для оставшихся 456 растений получили GLMM, считая долю посещений цветков одного экземпляра V. arvensis зависимой переменной, а прочие признаки — независимыми.
Проведенное разностороннее исследование подтвердило гипотезу о том, что все четыре дикие популяции V. arvensis за последние десятилетия стали реже размножаться при участии насекомых и чаще использовать самоопыление — по оценкам авторов, в среднем на 27% чаще. Эксперименты с изучением предпочтений шмелей показали, что те охотнее посещают цветки «воскрешенных» растений, а не современных популяций. В соответствии с этим признаки цветков, которые привлекают опылителей (прежде всего, объем нектара), также значительно уменьшились — в среднем на 20%.
При этом многие другие признаки (относительное число аллелей, генетическое разнообразие) у изученных фиалок не имеют универсальных трендов. Их не удалось проследить и для тех морфологических признаков, которые не участвуют во взаимодействиях с опылителями (длина чашелистиков, характеристики вегетативных органов и общий размер растения).
Для прочих признаков цветков также отмечены статистически значимые различия между «предковыми» и «потомковыми» популяциями фиалок. Площадь цветка V. arvensis сократилась в среднем на 10%, что означает и пропорциональное уменьшение длины венчика и ширины губы. Анализ данных морфометрии подтвердил заметные различия между «воскрешенными» и современными популяциями, описанные при сравнении отдельных чисел — на графике LDA (рис. 3) они образуют два разных кластера. Также отмечено небольшое по величине, но статистически значимое увеличение способности V. arvensis продуцировать семена в отсутствие опылителей.
Помимо общих трендов, фиалки разных локаций отличались по отдельным признакам, которые соответствуют разнообразию их местообитаний. При этом общая тенденция перехода к маленькому, более невзрачному цветку с меньшим количеством нектара отмечена для всех четырех локаций, хотя они и различаются по выраженности эффекта. Авторы подчеркивают, что выявленные изменения цветков V. arvensis не связаны с инбредной депрессией, то есть снижением жизнеспособности организмов из-за близкородственного скрещивания. Согласно оценкам на основании морфологических признаков цветка, величина инбредной депрессии примерно равна 3%.

Стоит отметить, что для двух локаций (Коммени и Круи) частота самоопыления в «предковой» и в «потомковой» популяциях не имеет значительных различий. Это контрастирует с тем, что фиалки из Коммени демонстрируют максимальные изменения морфологии цветка. Авторы связывают такие различия с тем, что смена стратегии опыления у V. arvensis там могла начаться раньше. Это согласуется с данными об исчезновении насекомых-опылителей уже в 1970-х годах. В пользу предположения говорит и максимальная среди всех «предковых» частота самоопыления именно у популяции из Коммени.
Таким образом, в обсуждаемой статье показана быстрая эволюция «синдрома самоопыления» (selfing syndrome) в диких популяциях растений, которую авторы связывают с антропогенной нагрузкой на их родные экосистемы. Современная биосфера столкнулась со многими глобальными вызовами — разрушением биотопов, загрязнением окружающей среды (в том числе связанным с сельским хозяйством и пестицидами), распространением инвазивных видов и т. д. Одно из важных последствий — снижение численности и биоразнообразия насекомых-опылителей, которое могло стать причиной быстрых эволюционных изменений в популяциях диких фиалок Франции.
Разумеется, переключение V. arvensis на самоопыление было возможно благодаря изначальной возможности к нему растения, то есть его самосовместимостью (самофертильностью). Самоопыление в той или иной форме (с переносом пыльцы между разными цветками одного растения, внутри одного цветка, в том числе не раскрывшегося) описано у множества цветковых: других фиалок, бобовых, орхидей и т. д. Ему могут способствовать особые механизмы переноса пыльцы — например, как в случае Erysimum incanum (желтушник седой), который для этого поглаживает пестик тычинками (Желтушник седой самоопыляется, поглаживая тычинками пестик, «Элементы», 11.02.2019). В то же время многие другие цветковые самонесовместимы за счет различных «барьерных» механизмов, которые препятствуют самоопылению — это двудомность, дихогамия, геркогамия и т. д.
В условиях недостатка опылителей переключение растений со смешанной стратегией доставки пыльцы на самоопыление позволяет добиться «тактического» преимущества. Однако в дальнейшем оно может обернуться «стратегическим» проигрышем в борьбе за жизнеспособность. Постоянный инбридинг вызывает неуклонные генетические изменения: уменьшается эффективный размер популяции и усиливается дрейф генов. Переход к такой стратегии размножения может оказаться необратим, а сама она — тупиковой, поскольку самооплодотворяющиеся популяции, возможно, имеют сниженную способность к адаптации и чаще вымирают.
Сложно сказать, сколько цветковых на фоне дефицита опылителей последовали примеру V. arvensis и стали чаще прибегать к самоопылению. Как бы то ни было, из-за резких эволюционных изменений в будущем они могут быстро потерять способность использовать насекомых для опыления из-за изменений в морфологии цветка, в том числе снижения объема нектара — очень «дорогостоящей» специализированной адаптации, которая служит только для привлечения насекомых. Может сработать принцип эко-эволюционного «порочного круга» (положительной обратной связи): исчезновение насекомых-опылителей и утрата энтомофильных адаптаций цветковых начнут все больше усиливать друг друга. Подобные изменения могут принять характер каскада и иметь опасные последствия для многих экосистем.
Исчтоник: Samson Acoca-Pidolle, Perrine Gauthier, Louis Devresse, Antoine Deverge Merdrignac, Virginie Pons, Pierre-Olivier Cheptou. Ongoing convergent evolution of a selfing syndrome threatens plant–pollinator interactions // New Phytologist. 2023. DOI: 10.1111/nph.19422.
Михаил Орлов
Свежие комментарии