
Растения — существа прикрепленные, они вынуждены изыскивать средства для распространения своего потомства (семян и спор). Лучший способ — пустить их по ветру; еще можно «заставить» подвижных животных разнести семена подальше или же, в крайнем случае, самостоятельно отбросить их как можно дальше. Последний вариант называют активным сбросом. Если для высших растений активный сброс семян — это интересное, но не слишком распространенное явление, то в микромире грибов и мхов это необходимость: без активного сброса споры не смогут вылететь за пределы зоны застойного воздуха или застрянут в пятне собственной слизи, или — в случае патогенов — останутся на уже зараженной поверхности. Спорам же нужно добраться туда, где господствуют воздушные потоки. Для достижения этой цели растения и грибы изобретательно используют различные физические силы и явления. В свежем выпуске журнала Current Biology американские и немецкие ботаники опубликовали два интересных обзора по этой теме — о них и пойдет речь ниже.
В свежем выпуске журнала Current Biology опубликованы два обзора, посвященные «баллистике» в растительном мире. Один из них написан Николасом Мани (Nicholas P. Money) из Университета Майями, США, другой — Аурелией Эмоне (Aurélia Emonet) и Анжелой Хей (Angela Hay) из Института растениеводства в Кельне, Германия. Авторы описывают, как различные растения и грибы стреляют спорами и семенами, достигая при этом замечательных результатов.
У растений и грибов отлажено множество способов рассредоточить свои споры и семена вокруг родительского растения.
Они пользуются ветром, животными-переносчиками или же применяют так называемый активный сброс. Активный сброс семян и спор помогает преодолеть зону застоя воздуха вблизи листьев и стеблей и достичь воздушных потоков, вылететь за пределы слизевого покрытия или зоны заражения, что особенно важно для грибов и патогенов. Для выстрела растению нужно преодолевать силу тяготения и силу трения о воздух. У высших растений семена достаточно тяжелые, поэтому они в основном «борются» с силой тяготения. А вот в микромире низших растений размеры «снарядов» маленькие и основным фактором становится трение о воздух. И у высших, и у низших растений выявлено большое разнообразие механизмов «стрельбы», достаточно мощных, чтобы преодолеть все эти трудности.Грибы и мхи оказались весьма изобретательны в использовании физики окружающего мира. Авторы приводят несколько досконально разобранных примеров этого разнообразия.
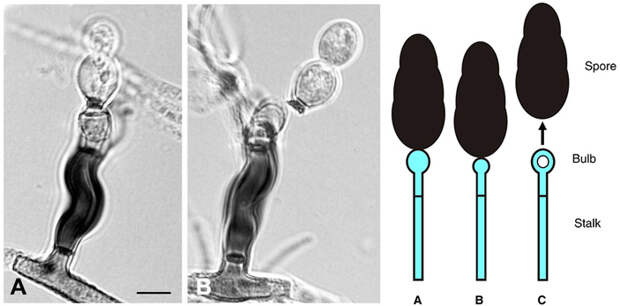
Гриб Deightoniella приспособился рассеивать свои споры за счет кавитации (ударной силы пузырьков в жидкости), а другой патогенный гриб Zygophiala присоединяет к кавитации еще и силу упругости. У дейтониеллы споры сидят на столбике конидиеносца. При испарении воды из клетки кондидиеносца давление в цитоплазме уменьшается — в результате формируется кавитационный пузырек. Этот пузырек разряжает энергию, ударяя в макушку расширенной клетки конидиеносца. Этот резкий удар придает ускорение конидии со спорами. Она отрывается и улетает на полмиллиметра в сторону, это примерно 15 длин споры.
Зигофиала обычно имеет искривленный конидиеносец. Он изгибается при испарении воды, в это же время формируются и кавитационные пузырьки. Они ударяют в столбик конидиеносца, и он резко, как пружина, распрямляется, так что конидий вылетает под действием импульса кавитационных пузырьков и выпрямления упругого стебелька конидиеносца.
На сравнительно большое расстояние (5–20 см) выстреливает спорами знакомый всем мох сфагнум. На этой высоте споры уже подхватываются воздушными потоками, выполняющими для мха задачу расселения. Но ключевой момент расселения — запустить споры выше застойного воздушного пласта. Как мох это делает? На концах коротких стебельков спорангиеносцев формируются спорангии круглой формы. Нижняя часть спорангия представляет собой воздушную камеру, а в верхней находятся споры. В стенках верхней части имеются псевдоустьица, через которые идет испарение влаги, в нижней, воздушной, части устьиц нет, она герметичная. При высыхании, а это происходит за несколько минут в хорошую погоду, спорангий сморщивается, приобретая форму цилиндра. В результате в воздушной камере увеличивается давление, достигая примерно 5 ат. Тогда крышечку спорангия срывает, и давление нижней камеры с силой выталкивает споры наружу вверх; при этом слышен хлопок, как при выстреле. Но этого мало: дополнительное ускорение придает спорам эффект вихревого кольца. Вихревое кольцо формируется при взаимодействии вылетающего под давлением воздуха из обрамленного валиком узкого горлышка спорангия (таким же образом курильщики пускают кольца дыма). Поэтому сфагнум использует для выстрела два механизма: давление сжатого воздуха и вихревое кольцо.

Еще мощнее стреляет папоротник. Он, как и гриб зигофиала, использует кавитацию и силу упругости. Работает это у папоротника следующим образом. На коротком стебельке развивается спорангий, все устройство которого обслуживает его артиллерийские нужды. Наружная часть спорангия представляет собой гребень из 12–13 клеток с плотными радиальными стенками и относительно гибкими латеральными. Это так называемый аннулюс. С другой стороны от аннулюса расположены тонкостенные клетки со щелью-стомиумом. Внутри головки спорангия находятся споры.
Все начинается с испарения воды из клеток аннулюса. Тогда их объем уменьшается, при этом сморщиваются мягкие латеральные стенки, а жесткие радиальные держат постоянную длину клеток (принцип аккордеона). В результате аннулюс отгибается назад, как пружина, стомиум раскрывается и споры повисают на остатках внутренних клеток головки. Дальнейшее испарение воды приводит к еще большему снижению давления внутри клеток аннулюса, в них образуются кавитационные пузырьки. Они сбрасывают энергию, ударяя в стенки клеток. Взведенный аннулюс резко распрямляется, и споры катапультируются. Но в катапульте есть еще один необходимый элемент — перекладина. Чтобы запустить ядро, а не просто уронить его на землю, рычаг катапульты должен удариться о перекладину — в момент удара ядру придается необходимый импульс. У спорангия папоротника такой перекладины нет, а ее функцию выполняет резкий переход от одного типа движения в другой. Сначала аннулюс разгибается под действием силы упругости — это очень быстрое движение, затем движение перехватывают пороэластичные силы, рассеивающие остаток энергии — это медленный процесс. На границе действия этих сил — замена перекладины — спора вылетает.

Хорошо нам знакомые грибы вешенки для запуска своих спор задействуют совсем другую силу, а именно силу поверхностного натяжения. Когда споры вешенки созревают, то на месте прикрепления споры к ножке (стеригме) формируется пробка с канальцем. Эта пробка напитывает влагу из воздуха, и она в какой-то момент начинает выделяться через каналец наружу особой каплей — так называемой каплей Буллера. В состав этой капли входит спирт маннит, обладающий свойством понижать точку росы и за счет этого конденсировать влагу вокруг себя. Поэтому нет ничего удивительного, что капля Буллера постепенно растет. Более того, на поверхности споры со стороны капли также синтезируется маннит, вместе с ним выносятся на поверхность неорганические ионы, которые совместно конденсируют дополнительную влагу над каплей Буллера.
И форма споры, и пробка на стеригме, и синтез маннита в точно выверенных местах нужны грибу, чтобы подготовиться к выстрелу. Он происходит следующим образом. Верхние капельки или капля стекают вниз к капле Буллера и сливаются с ней. Они встречаются ровно в месте прикрепления споры к ножке. В момент слияния капелек центр тяжести споры смещается, и она отрывается от ножки. Одновременно из-за слияния мелких капелек в одну большую высвобождается избыточная энергия сил поверхностного натяжения: поверхностное натяжение одной большой капли меньше суммы соответствующих сил нескольких мелких капель. Этой энергии оказывается достаточно, чтобы придать споре хорошее ускорение. Спора при этом начинает двигаться в направлении, противоположном стеканию капли и вдоль линии поверхности споры, по которой стекает капля (F. Liu et al., 2017. Asymmetric drop coalescence launches fungal ballistospores with directionality). Топология поверхности споры, таким образом, определяет направление прыжка споры наружу. А там споры уже подхватывает ветер. Кроме вешенки подобным образом выстреливают свои споры лисички, сыроежки и маслята. Пробка на месте прикрепления споры с канальцем, синтез маннита, форма споры — эти составляющие грибы приспособили и пустили в ход для нужд своей «артиллерии». Движущей силой выстрела они выбрали силу поверхностного натяжения.

Также в ходу у стреляющих грибов использование тургорного давления. Манипуляция тургорным давлением — обычное для растений дело, приспособили они его и для стрельбы. Например, этот механизм используют многие аскомицеты для вызгрузки своих спор. У аскомицета Pilobolus спорангиеносец представляет собой стебелек в несколько мм высотой, на конце которого имеется расширение, к которому прикреплен спорангий. Все три элемента важны: в стебельке синтезируются маннит и другие низкомолекулярные спирты, увеличивающие осмотическое давление в стебельке; раздутая верхняя клетка играет роль линзы, так что стебелек поворачивается к свету (это своего рода «глаз» гриба); а сверху сидит спорангий со спорами. Когда давление вырастает до 5 ат, раздутая клетка лопается, и спорангий стартует со скоростью 9 м/с (32 км/ч) — он отлетает на расстояние до 2,5 м!
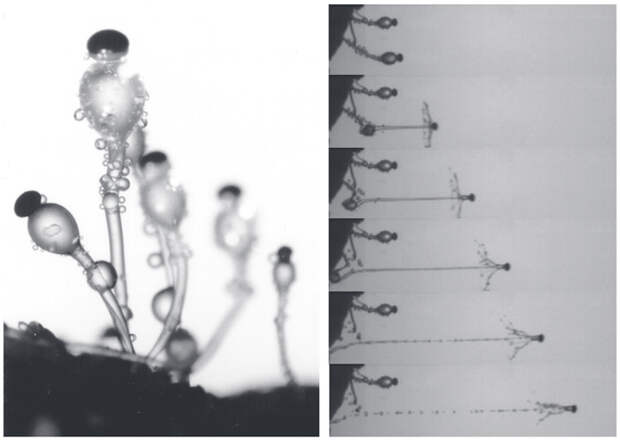
Авторы предполагают, что синтез маннита и других низкомолекулярных спиртов в спорангиеносцах аскомицетов могли унаследовать базидиомицеты, которые подобно вешенкам используют его для понижения точки росы на спорах. В обоих случаях маннит задействован для активной выгрузки спор, но работают при этом разные силовые механизмы.
Высшие растения для запуска своих семян используют те же физические силы — повышенное тургорное давление и силу упругости. Их действие хорошо иллюстрируется на примере сердечника шершавого. Механизм стрельбы у этого вида изучен на всех уровнях организации — от целого растения и тканей до молекулярного строения и генетики.

Сердечник шершавый (Cardamine hirsuta) — растение средней полосы. На созревших растениях формируются длинные тонкие стручки, которые, как хорошо всем известно, чуть тронешь — выстреливают семенами.
Все виды сердечников обладают этой способностью, потому их в просторечье называют недотрогами. Система запуска семян у них более или менее одинакова. Две половинки линейного стручка соединены друг с другом; по мере их роста напряжение в верхнем слое клеток створок (экзокарпе) нарастает, а во внутреннем (эндокарпе) — остается неизменным. Когда напряжение достигает максимума (к моменту созревания семян), достаточно малейшего толчка, чтобы створки оторвались друг от друга и скрутились (см. видео из обсуждаемой статьи A. Emonet и A. Hay). Это движение очень быстрое, его импульс и запускает семена, прикрепленные к пластинке внутри стручка.
Сердечнику шершавому даже не обязательно ждать высыхания стручков, чтобы выстрелить семенами. Его стручки могут скручиваться, и весьма эффективно, и без сухого растрескивания. Для этого у него выработались особые приспособления.
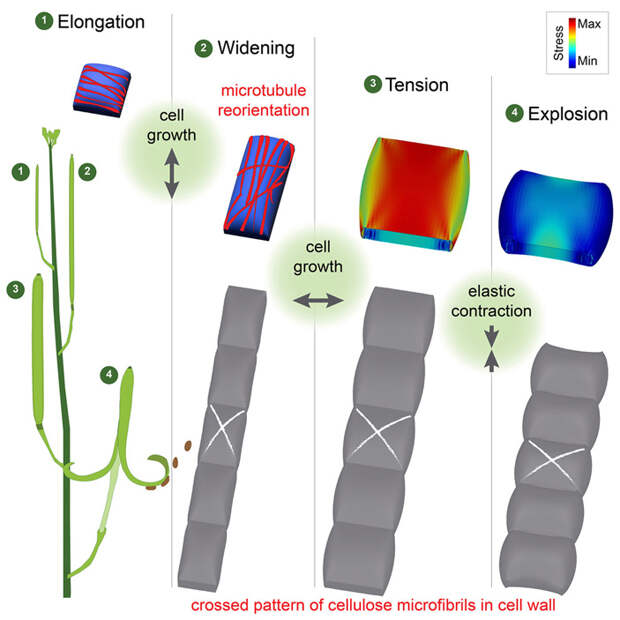
В слое экзокарпия стручков накапливается большое напряжения в силу увеличения тургорного давления. Но клетка умеет управлять этим давлением, распределяя его в нужном ей направлении. Оно организовывает свой рост так, чтобы слой экзокарпия стремился уменьшиться в длину и, соответственно, расшириться. Для этого клетки экзокарпия видоизменили геометрию своего роста. Они стали расти вширь, а не в длину. Подсчитано и доказано на модели, что в расширенной клетке напряжение увеличивается сильнее, чем в удлиненной (G. Mosca et al., 2024. Growth and tension in explosive fruit). Ученые пронаблюдали, что на начальных стадиях развития стручка клетки экзокарпия растут в длину, но ближе к концу созревания они начинают расти в ширину. Это происходит за счет изменения роста микрофибрилл целлюлозы: если раньше они росли перпендикулярно длине, не мешая клеткам удлиняться, то потом они резко поворачивают рост на 90°. Теперь клетке с вертикальными ребрами жесткости удобнее расширяться. Клетка проделывает этот трюк, изменяя доминирующее направление микротрубочек цитоскелета. Известно, что фибриллы целлюлозы строятся по микротрубочкам, так что при смене ориентации микротрубочек, перенаправляется и рост целлюлозных фибрилл.
Более того, появляются фибриллы, которые расположены к горизонтальным фибриллам под углом около 14°. Они, во-первых, ограничивают расширение клеток до положенных им 50 мкм, а, во-вторых, еще больше увеличивают напряжение в слое клеток. Экзокарпий в результате приобретает изгиб наружной поверхности и заряжается упругой энергией. Максимальное значение энергии в центре клеток, оно уменьшается к краям, поэтому в расслабленном состоянии клетка стремится укоротиться и расшириться.
В заряженном состоянии экзокарпий удерживается внутренним слоем клеток (эндокарпием). Так как у сердечника шершавого в экзокарпии накапливается сравнительно (по сравнению с другими сердечниками) высокое напряжение, то и эндокарпий должен обладать повышенной жесткостью, иначе растянутые клетки экзокарпа его разорвут, не дожидаясь созревания семян. Для увеличения прочности в клетках экзокарпия откладывается лигнин. Причем его отложение сосредоточено в той части клетки, которая обращена внутрь стручка, а прилегающая к экзокарпию сторона остается эластичной. Синтез лигнина направляется тремя ферментами, ответственными за удлинение его цепочек. Их работа зависит от присутствия меди, поэтому задача растения — накопить к моменту созревания побольше меди.
Лигнин в клетках эндокарпа отлагается не просто сплошной пластиной, а приобретает специфический облик трехчастной бляшки. Она сформирована таким образом, что разрешает растяжение вширь верхней части клетки, при этом удерживая определенный размер в нижней части.

Взрыв стручка, за которым следует выброс семян, ради которого все и устраивалось, напоминает по своей механике самоскручивающийся браслет. У такой системы два устойчивых состояния, причем из одного из них система легко переходит в другое. Эта система определяется геометрией и свойствами наложенных слоев. В створках стручка скреплены два слоя — верхний немного выпуклый, с накопленным напряжением в центре, и нижний, плоский. Первое устойчивое состояние — строго выпрямленное, нижний слой удерживает краями верхний. Выпрямление обеспечивается слоем эндокарпия с его особыми клетками, способными расширяться и жестко удерживать это состояние с помощью трехчастной лигниновой бляшки. Если хоть чуточку нарушить плоскость верхнего слоя, то система мгновенно переходит во второе устойчивое состояние: резко скручивается, снимая тем самым напряжение верхнего слоя. Вот таким образом стручок выстреливает семенами.
Эти работы показывают, как разнообразно устроено движение у растений, этих изначально неподвижных организмов. Как мы увидели, они используют буквально все силы, чтобы преодолеть ограниченность своего пространства.
Источники:
1) Nicholas P. Money. Microballistics in fungi and plants // Current Biology. 2024. DOI: 10.1016/j.cub.2024.06.027.
2) Emonet Aurélia, Angela Hay. Explosive seed dispersal // Current Biology. 2024. DOI: 10.1016/j.cub.2024.07.050.
Елена Наймарк
Свежие комментарии