
Какое отношение друг к другу имеют конечности разных животных? Есть ли у них что-то общее со щупальцами, которыми добывают пищу разные беспозвоночные? А крылья насекомых — это конечности или нет? Академик Владимир Васильевич Малахов и его коллега Михаил Маркович Ганцевич опубликовали статью о происхождении и путях эволюции двусторонне-симметричных животных (билатерий), в которой особое внимание уделено проблеме происхождения конечностей. Приведены доводы за то, что конечности членистоногих (и, судя по современным данным — даже крылья насекомых) произошли в конечном счете от щупалец далекого предка билатерий, который был похож на кораллового полипа.
Происхождение билатерий
Билатерии — крупнейшая ветвь животного царства, куда входят всевозможные двусторонне-симметричные животные от червей до позвоночных. Обсуждение происхождения той или иной группы живых организмов логично начать с вопроса, кто ее ближайшие родственники. Ближайшие родственники билатерий — это стрекающие, разнообразная группа животных, которая делится на две ветви: Anthozoa (коралловые полипы) и Medusozoa (сцифоиды, гидроиды и кубомедузы). Главное отличие между ними — то, что в жизненном цикле Medusozoa есть стадия медузы. В некоторых случаях она может исчезать (например, у пресноводной гидры), но у кораллов она отсутствует первично: они просто «не изобрели» ее. Кораллы — более древняя и примитивная ветвь стрекающих. Это доказывается, например, тем, что только у кораллов митохондриальная ДНК (как известно, унаследованная эукариотами от бактерий) остается кольцевой, как обычная бактериальная ДНК. А вот у Medusozoa митохондриальная ДНК стала линейной — это ярко выраженная производная черта (D.
Bridge et al., 1992. Class-level relationships in the phylum Cnidaria: evidence from mitochondrial genome structure). Кораллы находятся ближе всех к основанию эволюционного древа стрекающих, а значит, и к общим предкам стрекающих с билатериями.Все коралловые полипы — донные животные. Плавать в водной толще могут только их личинки. Кроме того, кораллы бывают одиночными или колониальными. Очевидно, что сравнивать с билатериями нужно одиночных кораллов, а не колониальных строителей рифов. У некоторых одиночных кораллов — например, у цериантарий — личинка не только хорошо плавает, но и питается другими планктонными организмами, то есть ведет планктотрофный образ жизни (рис. 1). Такой жизненный цикл свойствен очень многим морским беспозвоночным: взрослая форма живет на дне, а личинка — в планктоне, перемещаясь на большие расстояния и обеспечивая расселение. Вероятно, такой жизненный цикл был и у предков билатерий.
Стрекающих по традиции считают радиально-симметричными животными, но на самом деле у коралловых полипов симметрия двусторонняя (см. Двусторонняя симметрия кораллов древнее радиальной, «Элементы», 26.10.2016). Это их общий признак с билатериями. Дело в том, что значительную часть тела у кораллов занимает сплющенная глотка, в которую ведет щелевидный рот. Плоскость глотки и служит плоскостью двусторонней симметрии: она делит тело полипа на две зеркально симметричные половины. У большинства Medusozoa двусторонняя симметрия потеряна, очевидно, в результате приспособления к сидячему образу жизни, который характерен почти для всех стрекающих, не считая медуз. Но тогда откуда она взялась у коралловых полипов? Двусторонняя симметрия — признак подвижных животных, активно ползающих по дну. Напрашивается предположение, что коралловые полипы произошли от подвижных предков.
Коралловые полипы — самая примитивная группа стрекающих. Значит, стрекающие произошли от животных, которые были донными (с планктонной личинкой), ползающими и двусторонне-симметричными (рис. 2). Радиальная симметрия стрекающих вторична. Единственные истинно (первично) радиально-симметричные животные — это гребневики (см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015). Раньше многие биологи считали гребневиков предками билатерий, но молекулярная филогенетика показала, что это неверно. Гребневики — совершенно иная ветвь животного царства. Что же касается общих предков стрекающих и билатерий, то они, судя по всему, по части признаков были больше похожи на билатерий, чем на стрекающих.

Главная особенность стрекающих, которая не встречается больше ни у кого, — стрекательные клетки (книдоциты), поражающие добычу ядом. Это сделало их очень эффективными хищниками и позволило перейти к образу жизни хищников-засадчиков, почти полностью отказавшись от подвижности. Стрекательные клетки — настолько уникальное и ценное изобретение, что хищные моллюски и некоторые другие билатерии иногда «воруют» их у съеденных стрекающих, встраивают в свои покровы и используют в собственных целях (см. Клептокниды).
Еще одна важная черта стрекающих — щупальца, окружающие рот. Упомянутые выше цериантарии, крупные одиночные коралловые полипы, входящие в относительно примитивную группу шестилучевых кораллов, имеют два круга щупалец: лабиальный (внутренний) и маргинальный (внешний). Другие кораллы сохраняют только маргинальный круг щупалец. С другой стороны, оба круга щупалец есть у некоторых гидроидных полипов (семейства Tubulariidae и Corymorphidae). У сцифоидных медуз маргинальный круг состоит из нитевидных щупалец, которыми медуза ловит добычу, а лабиальный, судя по всему, преобразовался в ресничные лопасти, окружающие рот и отправляющие уже пойманную добычу внутрь (V. V. Malakhov, 2016. Symmetry and the tentacular apparatus in Cnidaria). Таким образом, система из двух кругов щупалец встречается и у кораллов, и у гидроидов, и у сцифоидов. Это дает основание полагать, что она уже была у общего предка всех стрекающих, а может быть, и у общего предка стрекающих с билатериями.
Метамерная теория
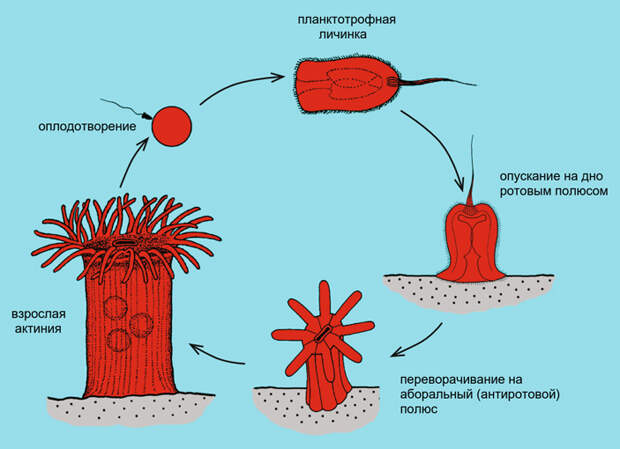
Как мог выглядеть подвижный общий предок стрекающих и билатерий? Если у него (подобно кораллам) был щелевидный рот, то он должен был ползать ротовой поверхностью вниз, собирая пищу со дна. Как он ползал? Скорее всего, ротовая (брюшная) поверхность была покрыта мерцательным эпителием, бьющиеся реснички которого постепенно двигали животное вперед. Так и сейчас ползают ресничные черви и многие моллюски. Ресничная подошва соответствовала лабиальному кругу щупалец, служа не только для отправки пищи в рот, но и для передвижения. Получается существо, похожее на перевернутого кораллового полипа. Интересно, что у некоторых настоящих коралловых полипов — актиний — планктонная личинка опускается на дно сначала ротовым полюсом, какое-то время ползает на нем и только потом переворачивается и прикрепляется к грунту, обращая рот вверх (рис. 3; см. также рис. 24 в статье J. F. Gemmil, 1920. The development of the sea anemones, Metridium dianthus and Adamsia palliata). Возможно, это рекапитуляция эволюционного пути, пройденного когда-то далекими предками. Во всяком случае, какое-то другое объяснение такого развития личинки придумать сложно.
Как был устроен самый первый представитель билатерий? На этот вопрос есть много разных ответов — от гипотез «простого предка», похожего на бескишечных ресничных червей (см. картинку дня Бескишечный червь), до гипотез «сложного предка», который должен был уже иметь сквозной кишечник (со ртом и анальным отверстием), центральную нервную систему, целом, метамерию и конечности. Наличие целома и метамерии во всех трех главных эволюционных стволах билатерий — у вторичноротых, линяющих и спиральнодробящихся — говорит в пользу гипотезы «сложного предка» (см., например: G. Balavoine, A. Adoutte, 2001. The segmented Urbilateria: a testable scenario).
Этой гипотезе соответствует метамерная теория происхождения билатерий, впервые предложенная в 1884 году Адамом Седжвиком (Adam Sedgwick; см. A. Sedgwick, 1884. On the origin of metameric segmentation and some other morphological questions). Изучая эмбриональное развитие онихофор (родственников членистоногих, которых иногда называют «бархатными червями»), Седжвик обратил внимание, что на определенной стадии этого развития бластопор (эмбриональный первичный рот), щелевидный и вытянутый вдоль брюшной стороны будущего тела, выглядит удивительно похоже на щелевидный рот коралловых полипов. Что, если это не случайное сходство? Сопоставив множество фактов, Седжвик предложил совершенно новую по тем временам концепцию эволюции животного царства.
Несколько раньше Рэй Ланкестер (Ray Lankester; E. R. Lankester, 1873. On the primitive cell-layers of the embryo as the basis of genealogical classification of animals, and on the origin of vascular and lymph systems) разделил многоклеточных животных на двуслойных (Diploblastica) и трехслойных (Triploblastica). У двуслойных животных гаструла состоит только из двух слоев клеток. У трехслойных добавился третий слой — мезодерма, из которой образуются мышцы, внутренний скелет, кровеносная система, стенки целомических полостей и выделительные органы. К трехслойным обычно относят только билатерий. Было естественно предположить, что от двуслойных предков сначала произошли какие-то очень просто устроенные билатерии (вроде плоских червей), потомки которых, постепенно усложняясь, приобрели целом и метамерию. Именно так выглядит, например, знаменитое эволюционное древо Эрнста Геккеля.
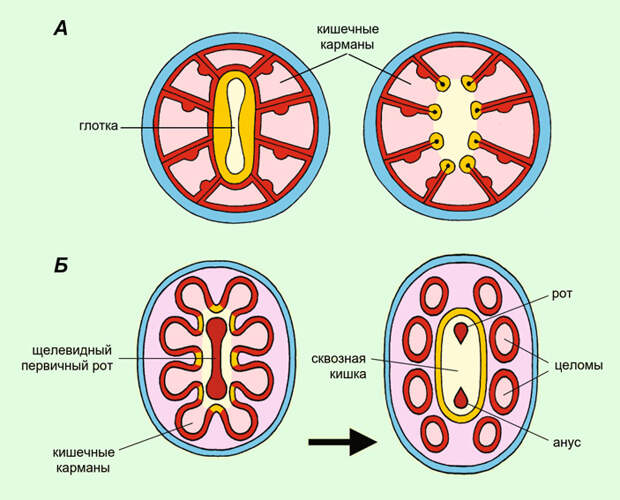
Седжвик же считал, что целомические метамерные билатерии произошли напрямую от двуслойного радиально-симметричного существа, которое было общим предком билатерий и стрекающих (рис. 4). Больше всего это существо напоминало кораллового полипа, начавшего ползать на ротовой стороне. В результате ползанья тело вытянулось, а двусторонняя симметрия усилилась. Щелевидный первичный рот тоже сильно вытянулся и закрылся посредине. Из его переднего и заднего концов образовались рот и анус билатерий. Так появился сквозной кишечник. Нервное кольцо, окружавшее первичный рот (и сохранившееся у стрекающих), превратилось у билатерий в брюшную нервную цепочку. Кроме того, у крупных коралловых полипов кишечник образует карманообразные выросты, отделенные друг от друга перегородками (септами). Это важно для увеличения поверхности, через которую могут всасываться питательные вещества. В момент возникновения билатерий эти кишечные карманы оказались на правой и левой сторонах, отделились от кишки и стали целомическими полостями. Поскольку их было несколько пар, то тем самым возникла и метамерия. Наконец, щупальца, находившиеся на краю ротового диска, у стрекающих так и остались щупальцами, а у билатерий превратились в конечности — параподии кольчатых червей, ножки онихофор и членистоногих.
Седжвик очень изящно вывел сложную конструкцию организма высокоорганизованных билатерий из предельно простых предпосылок. Но насколько этот сценарий соответствует реальному ходу эволюции?
Генетика развития и палеонтология
Современная генетика развития показывает, что у кораллов и у билатерий есть система экспрессии генов семейства Hox, которые один за другим «включаются» вдоль тела, деля его на физиологически различные области (см. Развитие «сегментов» у книдарий контролируется Hox-генами, как у билатерий, «Элементы», 01.10.2018). Экспрессия Hox-генов при этом образует характерный ступенчатый рисунок. У губок и гребневиков такой системы нет, а вот у кораллов и билатерий она, как выяснилось, выглядит очень похоже. Но у билатерий экспрессия Hox-генов разворачивается вдоль продольной оси тела, а у коралловых полипов — в плоскости глотки. Например, разные Hox-гены у них экспрессируются в разных парах кишечных карманов (см. рис. 1 в упомянутой статье на «Элементах»). Несомненно, что «с точки зрения» Hox-генов глоточная плоскость кораллов соответствует срединной (сагиттальной) плоскости билатерий. Это сильный довод в пользу метамерной теории.
Более того, недавно появилась гипотеза, что у общего предка стрекающих и билатерий было гораздо больше Hox-генов, чем у современных кораллов (B. M. Steinworth et al., 2023. Gene loss may have shaped the cnidarian and bilaterian Hox and ParaHox complement). После разделения ветвей билатерий и стрекающих и в той, и в другой ветви некоторые Hox-гены исчезли. Особенно много их исчезло в группе Medusozoa, которая почти утратила остатки двусторонней симметрии, так что сложная регуляция развития Hox-генами стала не нужна.
Амфистомия, то есть образование сквозного кишечника путем разделения щелевидного бластопора на два отверстия — способ развития, встречающийся в большинстве крупных групп билатерий (кольчатые черви, членистоногие, онихофоры, моллюски и другие). Эти факты хорошо знал уже Седжвик. Генетика развития дополняет их данными о генах Brachyury, FoxA и Cdx (рис. 5). У билатерий эти гены имеют отношение к процессу гаструляции и к формированию мезодермы. Гены FoxA и Brachyury экспрессируются в средней части вытянутого бластопора, а у хордовых — в хорде (см. У предков билатерий уже была примитивная протохорда, «Элементы», 18.09.2014). Ген Cdx у билатерий всегда экспрессируется в самой задней части кишки, вокруг анального отверстия (см. Дупликация гомеобоксных генов могла быть одной из причин кембрийского взрыва, «Элементы», 13.02.2015). У коралловых полипов картина очень похожа: гены FoxA и Brachyury экспрессируются вокруг средней части щелевидного рта, а ген Cdx — у одного из его концов.
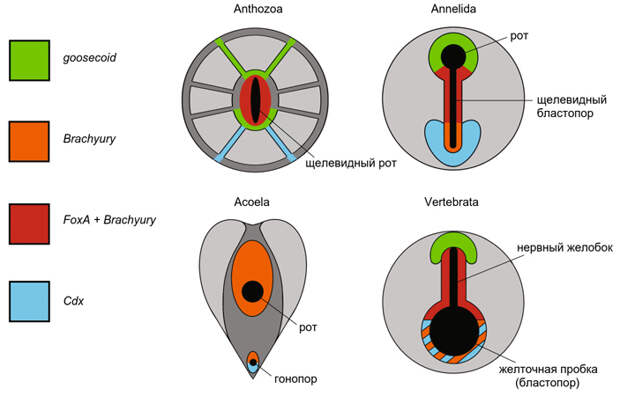
Интересно, что у бескишечных ресничных червей, которые не имеют не только сквозного, но и вообще никакого кишечника, ген Cdx экспрессируется вокруг отверстия половой системы — гонопора (см., например: A. Hejnol, J. M. Martín-Durán, 2015. Getting to the bottom of anal evolution). Не значит ли это, что отсутствие анального отверстия у бескишечных ресничных червей вторично? Возможно, у их предков одно и то же отверстие служило анусом и гонопором, а после редукции кишечника стало только гонопором.
Кишечная полость коралловых полипов четко делится на две части: центральный канал и карманы, предположительно соответствующие целомам билатерий (см. рис. 4). Эти карманы разделены мускулистыми септами. В клетках центрального канала кишки экспрессируются гены пищеварительных ферментов и регуляторных факторов, характерных для кишечника — тех же, что и у билатерий. А в кишечных карманах экспрессируются регуляторные гены, которые у билатерий маркируют целомическую выстилку, скелетные мышцы, сердце и другие производные мезодермы. Здесь генетика развития тоже подкрепляет метамерную теорию.
Метамерная теория косвенно подтверждается и палеонтологией. Крупные организмы позднего докембрия (эдиакария), которых иногда называют вендобионтами, в большинстве своем обладают отлично выраженной метамерией (рис. 6). О природе вендобионтов идут бесконечные споры, но последние данные определенно указывают на то, что они относятся к многоклеточным животным (см. Подтверждена принадлежность дикинсонии к животному царству, «Элементы», 24.09.2018). Возможно, у вендобионтов было даже две системы метамерных структур: упругое спинное «стеганое одеяло», придававшее животному постоянную форму, и «мягкое подбрюшье» — многочисленные правые и левые кишечные карманы, которые точно так же, как у кораллов, служили для транспорта и всасывания питательных веществ (см., например: J. Dzik, 2003. Anatomical information content in the Ediacaran fossils and their possible zoological affinities). При этом для вендобионтов чрезвычайно характерна симметрия скользящего отражения, когда метамеры правой и левой сторон расположены со сдвигом. Симметрия скользящего отражения почти не встречается у современных билатерий (одно из немногих исключений — ланцетник, у которого именно так расположены мышечные сегменты — миомеры). Но она очень легко формируется у коралловых полипов, например у тех же цериантарий. Дело в том, что новые септы у них образуются в зоне роста (находящейся на одном из концов вытянутой глотки) строго последовательно, одна за другой. Поэтому в каждый данный момент число септ и кишечных карманов, расположенных у полипа справа и слева от глотки, чаще всего не совпадает. И располагаются они со сдвигом. У вендобионтов новые правые и левые метамерные единицы тоже формировались одна за другой в зоне роста, находившейся у заднего конца тела (J. Dzik, 1999. Evolutionary origin of asymmetry in early metazoan animals). Так что источник сдвига симметрии тут общий.

И наконец, о щупальцах. Генетика развития выяснила, что формирование кишечных карманов и щупалец у кораллов контролируется одними и теми же Hox-генами. Выключение гена, «отвечающего» за развитие определенной пары кишечных карманов, приводит к тому, что связанные с этими карманами щупальца либо развиваются с нарушениями, либо отсутствуют вовсе. Примерно так же у членистоногих (например, у всем известной мухи-дрозофилы) мутации Hox-генов одновременно влияют на развитие как сегментов туловища, так и сидящих на них конечностей. Это довод в пользу того, что у общего предка билатерий со стрекающими уже были не только метамерные кишечные карманы, но и круги щупалец. Что же произошло с этими щупальцами дальше?
Ресничные шнуры
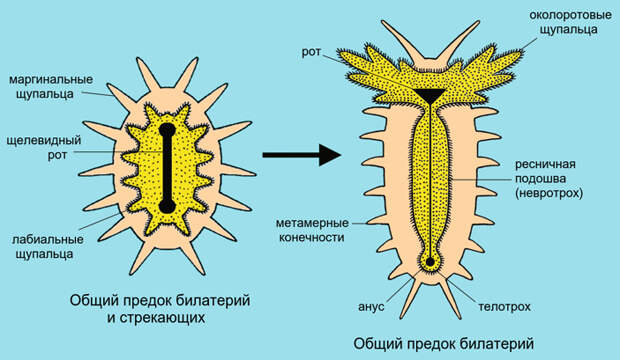
Когда гипотетический предок билатерий опустился на дно и начал ползать на ротовой стороне, лабиальный (внутренний) круг щупалец был вынужден взять на себя функцию ресничной подошвы, движущей животное вперед, и одновременно — ротового аппарата, направляющего в рот пищевые частицы. Когда вскоре после этого вытянутый первичный рот закрылся посредине, разделившись на передний рот и задний анус, бывший лабиальный круг щупалец тоже преобразовался. От него остались околоротовое и околоанальное ресничные поля, соединенные остатком ресничной подошвы (рис. 7). В самом полном виде эти элементы сохранились у некоторых планктонных личинок (см. Общее происхождение трохофор и диплеврул: за и против, «Элементы», 06.02.2020).
Сложнее всего устроено околоротовое (адоральное) ресничное поле. У планктотрофных личинок билатерий оно окаймлено ресничным шнуром, то есть полоской клеток с особо мощными ресничками (рис. 8). У ресничных личинок первичноротых — трохофор — разные отрезки этого ресничного шнура называются прототрохом (впереди рта) и метатрохом (позади рта). У трохофоры околоротовое ресничное поле работает по так называемому downstream-механизму: прототрох и метатрох бьют ресничками таким образом, чтобы гнать воду вместе с пищевыми частицами навстречу друг другу — в рот. Во взрослом состоянии у некоторых первичноротых, например у сидячих кольчатых червей, отрастают околоротовые щупальца, работающие в точности по тому же принципу: по периметру щупалец тянется ресничный шнур, который улавливает пищевые частицы и гонит их вместе с водой в сторону рта.

Кроме того, у типичной трохофоры есть телотрох — ресничный шнур, окружающий анальное отверстие и служащий исключительно для движения. Околоротовое и околоанальное ресничные поля соединены тонкой ресничной полосой — невротрохом (бывшая ресничная подошва). Прототрох, метатрох, невротрох и телотрох в сумме соответствуют древнему лабиальному кругу щупалец.
У вторичноротых околоротовой ресничный шнур называется неотрохом и состоит не из ресничных, а из жгутиковых клеток (это более примитивное состояние). У личинки полухордовых — торнарии — неотрох продолжается в узкий невротрох, а тот — в мощный телотрох, который охватывает анальное отверстие и служит для движения. Такой план расположения ресничных шнуров нетрудно сопоставить с планом расположения ресничных шнуров трохофоры. Неотрох, невротрох и телотрох в сумме соответствуют древнему лабиальному кругу щупалец. У иглокожих, которые во многих отношениях гораздо более специализированы, неотрох приобретает очень сложную форму, а невротрох и телотрох, наоборот, редуцируются.
Проблема в том, что у вторичноротых, в отличие от большинства первичноротых, захват пищи ресничными шнурами идет по так называемому upstream-механизму: жгутик гонит воду не в сторону рта, а в противоположную сторону, при этом извлекая из потока воды пищевые частицы и отбрасывая их ко рту обратным движением. У взрослых сидячих полухордовых — крыложаберных, которые фильтруют воду ресничными щупальцами, эти щупальца тоже работают по upstream-механизму. Впрочем, различие между первичноротыми и вторичноротыми тут не абсолютно. Есть и промежуточные случаи. У личинок форонид, которые, несомненно, относятся к первичноротым, но очень примитивны по ряду признаков (см. Предок мшанок был «протофоронидой», «Элементы», 03.04.2018), преоральный ресничный шнур, гомологичный прототроху (см. Е. Н. Темерева, В. В. Малахов, 2009. Развитие форонид), работает по downstream-механизму, а посторальный ресничный шнур, гомологичный метатроху — по upstream-механизму, как у вторичноротых. Ресничные щупальца, с помощью которых питается взрослая форонида, образуются только на посторальном ресничном шнуре и опять-таки захватывают пищу по upstream-механизму. Это подтверждает, что система ресничных шнуров первичноротых и вторичноротых может иметь общее происхождение.
Щупальца и конечности
Предположение, что щупальца кораллоподобного предка дали начало конечностям билатерий, с самого начала было частью метамерной теории. Особенно интересны в этом свете конечности линяющих животных — Ecdysozoa, которые устроены довольно сложно и расположены строго метамерно: каждая конечность принадлежит одному сегменту (в отличие от конечностей позвоночных).
Открытие эволюционной ветви Ecdysozoa стало одним из символов революции в зоологии, случившейся на рубеже XX–XXI веков (см. картинку дня Афротерии). В 1997 году вышла историческая статья, в которой эта ветвь была выделена на основании исследований рибосомной РНК (A. M. Aguinaldo et al., 1997. Evidence for a clade of nematodes, arthropods and other moulting animals). Надо сказать, что независимо от молекулярных филогенетиков к этому открытию вплотную подошли и палеонтологи. Еще в 1989 году Ежи Дзик (Jerzy Dzik) и Гюнтер Крумбигель (Günter Krumbiegel) опубликовали статью, содержавшую примерно те же выводы о происхождении членистоногих, которые потом стали общепринятыми благодаря «экдизозойной» гипотезе (J. Dzik, G. Krumbiegel, 1989. The oldest ‘onychophoran’ Xenusion: a link connecting phyla?). Главная особенность группы Ecdysozoa — кутикула, способная служить наружным скелетом. Чтобы животное могло беспрепятственно расти, кутикула периодически сбрасывается во время линьки, которую запускает стероидный гормон экдизон.
Кто же такие Ecdysozoa? В современной фауне к этой группе относятся членистоногие вместе с их более примитивными родственниками (онихофорами и тихоходками) и несколько групп червей: нематоды, приапулиды, киноринхи, лорициферы и волосатики. Экдизозойная кутикула несовместима с ресничками, поэтому никаких ресничных шнуров, ресничных подошв и ресничных щупалец у Ecdysozoa не бывает. Зато конечности у них появились очень рано. Некоторые палеонтологи считают, что червеобразные Ecdysozoa возникли в результате редукции конечностей, связанной с переходом к рытью в грунте (А. Ю. Журавлёв, 2014. Ранняя история Metazoa — взгляд палеонтолога). Если это верно, значит, уже у самых первых Ecdysozoa конечности были.
Откуда они взялись? Поиску ответа может помочь тот факт, что закладка рта и ануса из переднего и заднего концов вытянутого бластопора (амфистомия) распространена у Ecdysozoa очень широко. Этот способ развития описан не только у онихофор и разных групп членистоногих, но и у таких крайне специализированных существ, как нематоды (В. В. Малахов, 1986. Нематоды: строение, развитие, система и филогения). Как и говорил Седжвик, это похоже на рекапитуляцию эволюционного этапа щелевидного рта, подобного рту коралловых полипов. С другой стороны, давно известно, что у трилобитов (план строения которых очень примитивен для членистоногих) и у некоторых ракообразных на первых члениках всех конечностей, вплоть до самой задней пары, сидят жевательные отростки (гнатобазы). Современная палеонтология подтверждает, что эти отростки появились в самом начале эволюции членистоногих (J. Yang et al., 2018. Early Cambrian fuxianhuiids from China reveal origin of the gnathobasic protopodite in euarthropods). Но еще в середине XX века классик сравнительной анатомии беспозвоночных Владимир Николаевич Беклемишев писал по этому поводу: «Все конечности являются здесь ротовыми конечностями, хотя они и сидят вдоль уже замкнувшегося первичного рта, тянущегося в прототипе вдоль всей брюшной стороны» (рис. 9).

Наконец, у Ecdysozoa, во всяком случае у членистоногих и онихофор, есть целомы — пусть и редуцированные у взрослых форм, но явно унаследованные от предков. Если эти целомы произошли от кишечных карманов кораллоподобного предка, значит, им когда-то сопутствовали и щупальца.
Во что же эти щупальца превратились? Здесь опять может дать подсказку палеонтология. Ископаемых Ecdysozoa из кембрийского периода (родственников членистоногих, которых иногда называют «стволовыми членистоногими»; см. Палеонтологи уточнили время появления первых членистоногих, «Элементы», 23.08.2018) известно много. Они разнообразны и порой весьма причудливы. Самое интересное для нас сейчас — то, что у многих из них на каждом сегменте сидит по две пары придатков (см. картинку дня Керигмахела). На брюшной стороне, ближе к средней линии тела, находятся ножки, адаптированные для хождения по грунту и способные телескопически вытягиваться. А над ними, дальше от средней линии, сидят лезвиеподобные боковые лопасти, помогающие животному плавать. Эти лопасти — именно самостоятельные придатки, а не просто расширения спинных хитиновых щитков, как можно было бы подумать (см., например: G. Budd, 1996. The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group). Нижние ножки предложено называть аннулоподиями («кольчатые ноги»), а верхние лопасти — ламеллиподиями («пластинчатые ноги»). Вероятно, это и есть гомологи, соответственно, лабиального и маргинального кругов щупалец.
Такое сочетание плавательных и ходильных конечностей, очевидно, позволяло древним экдизозоям неплохо плавать — в основном вблизи дна, а при необходимости опускаться на дно и ходить по нему. У тех из них, кто специализировался в сторону плавания, ламеллиподии сильно разрослись, да еще и появились дополнительные плавательные лопасти, образующие хвостовой веер (рис. 10). Аннулоподии, наоборот, редуцировались. Именно так обстоит дело у «звезд» палеонтологии кембрия — загадочной пятиглазой опабинии (Opabinia) и крупного хищного аномалокариса (Anomalocaris; см., например: Найдено «недостающее звено» между членистоногими и загадочными кембрийскими чудовищами аномалокарисами, «Элементы», 16.02.2009). Интересно, что у некоторых особенно крупных — двухметровых! — родственников аномалокариса аннулоподии, вместо того чтобы редуцироваться, превратились в дополнительную пару ламеллиподий (см. Новооткрытый родственник аномалокариса проливает свет на происхождение конечностей членистоногих, «Элементы», 16.03.2015). Возможно, у них произошло что-то вроде гомеозисной мутации (см. Современные высшие растения возникли в результате сдвига экспрессии генов, «Элементы», 10.04.2018), которая при таком размере животного оказалась полезной. Напротив, у тех экдизозоев, кто специализировался к жизни на поверхности дна, редуцировались ламеллиподии, а аннулоподии усложнились и стали членистыми. Так появились членистоногие.
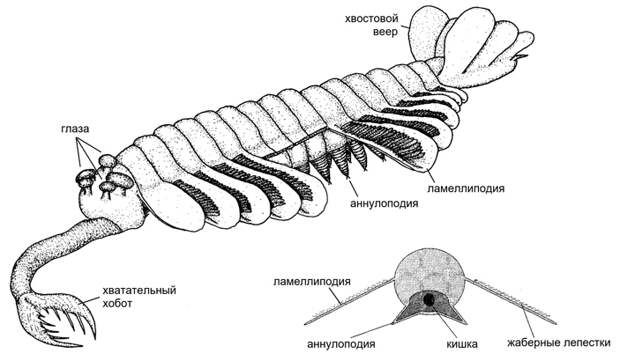
Двуветвистая конечность
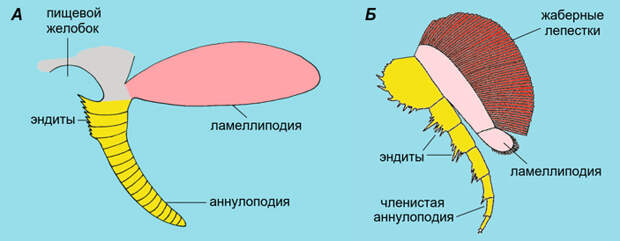
У членистоногих бывшая аннулоподия, ставшая теперь членистой ногой, и остаток ламеллиподии слились в единую двуветвистую конечность. Ламеллиподия у них редуцировалась не до конца. Дело в том, что у ламеллиподии с самого начала было две функции — плавательная и дыхательная. Например, у опабинии (которая к членистоногим не относится) хорошо видно, что на ламеллиподиях сидят жаберные лепестки. Вот эти жаберные лепестки от них в основном и остались. Они сформировали верхнюю ветвь конечности членистоногих, которая у ракообразных называется эпиподитом.
Первично двуветвистую конечность, служащую прототипом всех остальных конечностей членистоногих (кроме антенн), можно увидеть у трилобитов (рис. 11). Почти настолько же примитивны дожившие до наших дней мечехвосты, мало изменившиеся за последние 400 миллионов лет (см. У мечехвостов обнаружены свидетельства полногеномной дупликации, «Элементы», 27.11.2015). У современных мечехвостов бывшие ламеллиподии превращены либо в жабры, либо в генитальные крышечки, либо (на одном из сегментов) в особый рудиментарный вырост под названием флабеллум. Но у их древних (силурийских) родственников конечности полноценно двуветвистые, причем обе ветви членистые (D. Briggs et al., 2012. Silurian horseshoe crab illuminates the evolution of arthropod limbs). Это еще раз подтверждает, что двуветвистая конечность — общая древняя особенность членистоногих.
У ракообразных конечность сильно усложняется (рис. 12). Прежде всего, основание, на котором сидят и эпиподит, и членистая ходильная нога, у них само становится многочленным (обычно в нем три членика). Эпиподитов — листовидных жаберных придатков — при этом тоже иногда становится два или три. В этом случае древняя жабра, когда-то сидевшая на ламеллиподии, как бы распадается на отдельные эпиподиты. Но это еще не вся сложность. Ходильная нога (бывшая аннулоподия) у ракообразных, в свою очередь, расщепилась на две ветви, которые принято называть экзоподитом и эндоподитом. Это — вторичная двуветвистость, которая в итоге привела к образованию трехветвистой конечности.

Благодаря биологии развития сейчас известно, что вторичное расщепление конечностей ракообразных произошло в результате разделения зоны роста, находящейся на кончике главной оси ноги (C. Wolff, G. Scholtz, 2008. The clonal composition of biramous and uniramous arthropod limbs). При этом расщепилась надвое зона экспрессии гена Dll, который обычно маркирует у билатерий кончики конечностей (см. Конечности у членистоногих и кольчатых червей растут из общего корня, «Элементы», 20.11.2008). Все это случилось только в эволюционной ветви ракообразных. Их экзоподит и эндоподит вместе гомологичны древней аннулоподии. Таким образом, свойственное ракообразным расщепление конечности на экзоподит и эндоподит — вовсе не та двуветвистость, которая наблюдается у трилобитов и мечехвостов. Дополнительное усложнение конечностей ракообразных, скорее всего, связано с богатством их функций у этих водных животных: они могут служить для хождения по грунту, могут — для плавания, могут подгонять воду ко рту или к жабрам, могут нести жабры на себе, могут служить сенсорами и так далее. Экзоподит, например, предназначен в основном для плавания, а эндоподит — для хождения. У некоторых ракообразных, например у жаброногов (Anostraca) и у щитней (Notostraca), конечности становятся даже многоветвистыми (см. Н. А. Заренков, 1982. Членистоногие. Ракообразные. Ч. 1).
Экдизозоям, освоившим сушу, дополнительные ветви конечностей не нужны. У онихофор на туловище остаются только аннулоподии, ни во что не преобразованные (потому онихофор и не относят к членистоногим). У многоножек и насекомых — только ходильные ноги, хотя и разнообразно устроенные. Одно время онихофор, многоножек и насекомых объединяли в группу одноветвистых, оказавшуюся искусственной (см. картинку дня Керигмахела). У онихофор и многоножек остаток ламеллиподии, судя по всему, исчез без следа. А вот у насекомых его судьба оказалась совершенно особой.
Происхождение крыльев
Одним из важнейших событий не только в эволюции членистоногих, но и во всей истории жизни на Земле было появление крылатых насекомых. По времени оно приблизительно совпадает с выходом позвоночных на сушу (см. С. А. Ястребов, 2016. Жизнь на суше: расцвет, кризис, возрождение). Насекомые — единственные беспозвоночные, овладевшие активным полетом. Благодаря этому они достигли небывалого эволюционного успеха. Сейчас на Земле живет больше видов насекомых, чем всех остальных животных вместе взятых (см. Global biodiversity). Биологов давно интересовало, как именно насекомым удалось взлететь. Важнейшей частью этого вопроса, конечно, стал вопрос, откуда у насекомых взялись крылья: они ведь должны были развиться из какого-то другого органа. «Будь у древних сравнительно-анатомическая точка зрения, они не могли бы вообразить драконов с крыльями на спине», — писал Владимир Николаевич Беклемишев. Насекомые в этом смысле подобны драконам: их крылья уникальны.
Есть много теорий происхождения крыльев насекомых. Более того, их было много уже сто лет назад (см. G. Crampton, 1916. The phylogenetic origin and the nature of the wings of insects according to the paranotal theory). В XX веке господствующей была версия, согласно которой крылья образовались из боковых выростов спинных щитков туловища, которые сначала служили для планирования, а потом приобрели подвижность. Эти гипотетические выросты называют паранотальными. Паранотальной теории придерживался, например, тот же Беклемишев. Альтернативные идеи, связывающие происхождение крыльев либо с жабрами, либо с конечностями, существовали давно (они появились даже раньше паранотальной теории), но большой популярностью не пользовались.
В начале XXI века стало ясно, что насекомые произошли не от многоножек, как считало раньше большинство исследователей, а от ракообразных (более того, с точки зрения кладистики они и есть ракообразные, но в эти схоластические детали мы здесь вдаваться не будем). Как очень часто бывает в эволюционной биологии, идея происхождения насекомых от ракообразных не была полностью новой — она имела сторонников и в прошлом веке (G. Crampton, 1938. The interrelationships and lines of descent of living insects). Но закрыть этот вопрос смогла только молекулярная филогенетика (см., например: В. В. Алёшин и др., 2009. О положении насекомых в кладе Pancrustacea). Это несколько поменяло перспективу и усилило позиции тех, кто считал, что крылья произошли все-таки от жабр. Например, Ярмила Кукалова-Пек (Jarmila Kukalová-Peck) еще в «домолекулярную» эпоху отвергала паранотальную теорию и настаивала на гомологии крыльев насекомых жаберным пластинкам их водных предков (J. Kukalova-Peck, 1978. Origin and evolution of insect wings and their relation to metamorphosis, as documented by the fossil record).
Кроме того, генетика развития довольно быстро выявила группу «крыловых» генов, которые экспрессируются в крыльях насекомых и в эпиподитах ракообразных, но не экспрессируются в ходильных ногах (N. Shubin et al., 1997. Fossils, genes and the evolution of animal limbs). Дальнейшие исследования регуляторных генов показали, что профиль их экспрессии индивидуален не только для каждой ветви конечности, но и для каждого ее членика. В итоге удалось провести очень тщательное экспериментальное исследование, которое привело к простому и ясному выводу: крыло насекомых гомологично эпиподиту наиболее проксимального (ближайшего к туловищу) членика конечности ракообразных (см. подробный разбор этого исследования в статье Гипотеза о происхождении крыльев насекомых от боковых отростков ног ракообразных проверена на бокоплавах-мутантах, «Элементы», 14.12.2020; вывод представлен в этой статье на рис. 5). Если это не будет опровергнуто, то вопрос о происхождении крыльев, видимо, можно считать решенным.
Означает ли это, что насекомые полетели прямо из воды? Ведь если бы они прошли этап полностью наземной жизни, жаберные придатки успели бы исчезнуть. Действительно, похоже, что предки крылатых насекомых были водными или в лучшем случае амфибиотическими (см. Д. Е. Щербаков, 2018. Происхождение крылатых насекомых: из воды — в воздух). Раннепалеозойские водоемы имели крайне нестабильные очертания берегов (см. картинку дня Ризодус), и расселяться из водоема в водоем было лучше всего по воздуху. Первыми такой способ расселения освоили высшие растения: предполагается, что у них именно для этого появился воздушный спорофит, способный выбрасывать в воздух споры (см. Современные высшие растения возникли в результате сдвига экспрессии генов, «Элементы», 10.04.2018). Насекомые последовали за ними. Несколько итальянских биологов обосновали эту гипотезу еще сорок лет назад в интересной статье «О филогении крылатых насекомых» (P. Omodeo et al., 1980. On the phylogeny of pterygote insects). Крылатые формы насекомых возникли как относительно короткоживущие расселительные стадии. Мы до сих пор видим такую жизненную стратегию у поденок, которые по многим признакам очень примитивны (см. упомянутую статью Дмитрия Евгеньевича Щербакова).
А можно ли считать, что крылья насекомых гомологичны маргинальным щупальцам кораллов? В рамках метамерной теории происхождения билатерий — да, хотя гомология эта и частичная (по изложенной версии в крыло превратился лишь один из нескольких эпиподитов). В любом случае глубина преемственности, которая здесь намечается, впечатляет.
Самой эволюционной биологии тоже свойственна глубокая преемственность. Мы видим много примеров того, как гипотезы, высказанные биологами XIX или начала XX века, получают поддержку от молекулярной филогенетики и биологии развития начала XXI века. Конечно, изложенный здесь эволюционный сценарий может быть спорным, но на то он и сценарий, чтобы быть предметом обсуждения. Биология целостна. Такие классические натуралисты, как гениальный Фрэнсис Бальфур (Francis Maitland Balfour) и его ученик Адам Седжвик, Эрнст Геккель (Ernst Haeckel) и его ученики Оскар и Рихард Гертвиги (Oscar Hertwig, Richard Hertwig), — перечисление можно продолжать долго — сделали много открытий и высказали много идей, которые остаются актуальными в современном дискурсе и подлежат проверке современными методами.
Источник: V. V. Malakhov, M. M. Gantsevich. The origin and main trends in the evolution of bilaterally symmetrical animals // Paleontological Journal. 2022. DOI: 10.1134/S0031030122080044.
Сергей Ястребов
Свежие комментарии