
Брахиоподы — морские щупальцевые животные, живущие в ракушках, развитие которых интересует зоологов уже много лет. В последнее время шла дискуссия о том, образуется ли у них целомическая полость из выпячиваний первичной кишки (энтероцельным путем) или путем расхождения клеток плотного зачатка (шизоцельным путем). Новое исследование показало, что первое предположение оказалось верным. Энтероцельный способ развития целома — признак, общий у брахиопод с вторичноротыми и, возможно, унаследованный и теми, и другими от древнейших двусторонне-симметричных животных. Кроме того, целомическая мезодерма брахиопод развивается из двух источников — переднего (головного) и заднего. Этот древний признак известен у многих других двусторонне-симметричных животных, но у брахиопод он обнаружен впервые.
Брахиоподы (Brachiopoda) — тип морских беспозвоночных, насчитывающий около 400 современных видов. Это сидячие животные, внешне похожие на двустворчатых моллюсков (до середины XIX века их и считали моллюсками). Но на самом деле брахиоподы относятся не к моллюскам, а к щупальцевым животным, главной особенностью которых является лофофор — круговой или подковообразный венчик щупалец, окружающий рот (но не анус). Щупальца лофофора покрыты ресничками и служат аппаратом для отцеживания из воды мелких пищевых частиц. Они гидростатически поддерживаются заходящей в них целомической полостью. Тело брахиоподы целиком, за исключением прикрепительного стебелька, заключено внутри двустворчатой раковины, которая может быть прикреплена к камню, заякорена с помощью стебелька в мягком осадке, а может и свободно лежать на дне. Тело животного занимает примерно треть раковины, ее передние две трети заняты лофофором, который непрерывно фильтрует воду (рис. 1).
Створки раковины брахиопод расположены совершенно иначе, чем у двустворчатых моллюсков, с которыми их поначалу путали. Сходство между ними — поверхностное: это хороший пример эволюционной конвергенции, вызванной похожим образом жизни. У двустворчатых моллюсков правая и левая створки раковины охватывают тело с боков. У брахиопод створки долгое время называли спинной и брюшной. Но датский зоолог Клаус Нильсен (Claus Nielsen) — один из лучших современных зоологов беспозвоночных, работы которого не раз обсуждались на «Элементах», — показал, что на самом деле личинка брахиопод при метаморфозе буквально складывается пополам брюшной стороной внутрь (C. Nielsen, 1991. The development of the brachiopod Crania (Neocrania) anomala (O. F. Müller) and its phylogenetic significance). При этом брюшная сторона сильно укорачивается, а на спинной стороне возникают две створки раковины — передняя и задняя, которые и заключают в себе тело животного. Никакой брюшной створки у брахиопод нет, они обе спинные.
Вероятно, предки брахиопод были червями, имевшими на спине две защитные пластинки. При нападении хищника они сворачивались, подставляя врагу защищенную спину. В наши дни так ведут себя мокрицы (наземные ракообразные), хитоны (примитивные моллюски) и даже некоторые млекопитающие — ежи, броненосцы, панголины. Со временем свернутая форма тела стала у этих малоподвижных существ постоянной и породила план строения животного, сложенного пополам (рис. 2; см. также рис. 5 в статье Древнейшие предки кольчатых червей могли быть похожи на брахиопод, «Элементы», 26.02.2016).

Брахиоподы — группа, расцвет которой остался в далеком прошлом. Разнообразнее всего они были в палеозое. Но и вымирать они, судя по всему, не собираются. Брахиоподы — одна из немногих групп животных, которые прослеживаются в палеонтологической летописи без перерыва на протяжении всего последнего полумиллиарда лет, от эпохи кембрийского взрыва до современности. Раковины брахиопод прекрасно сохраняются, и палеонтологи очень любят их описывать.
Брахиоподы вместе с другими щупальцевыми животными принадлежат к эволюционной ветви спиральнодробящихся, или просто спиралий (Spiralia). Главный признак этой ветви фигурирует в ее названии. Дело в том, что у многоклеточных животных есть два основных способа дробления яйца — радиальное и спиральное (см. Существование эволюционной ветви линяющих животных получило эмбриологическое подтверждение, «Элементы», 23.09.2014). Радиальное дробление организовано проще, в этом случае зародыш долго сохраняет внешнюю радиальную симметрию. При спиральном дроблении возникающие клетки совершают закономерные повороты то по часовой стрелке, то против нее, образуя спиральные фигуры (см. рис. 2 в статье Платизои ставят под сомнение происхождение двусторонне-симметричных животных от целомического предка, «Элементы», 23.06.2014). Спиральное дробление устроено почти одинаково у таких непохожих друг на друга животных, как кольчатые черви и моллюски. Но вот у брахиопод дробление радиальное (S. Santagata, 2015. Brachiopoda). Или они почему-то утратили спиральность дробления, или, как считает академик Владимир Васильевич Малахов, их радиальное дробление — это сохранившийся архаичный признак. «На самом деле этот тип эмбриогенеза — древняя черта всех билатерально-симметричных животных; только одни из них ее сохранили, а другие — нет» (В. В. Малахов, 2009. Революция в зоологии: новая система билатерий).
Брахиоподы — целомические животные. Это означает, что у них есть вторичная, или целомическая, полость тела, образованная средним зародышевым листком — мезодермой. По сути дела, целом — это мешок из мезодермального эпителия, заполненный жидкостью. Принято считать, что у целома четыре основных функции: распределительная (примерно как у кровеносной системы), выделительная (в целом обычно открываются выделительные трубочки — метанефридии), половая (у брахиопод яйцеклетки и сперматозоиды выводятся наружу через те же метанефридии) и опорная — например, в щупальцах лофофора заходящий туда целом, как уже говорилось, служит гидростатическим скелетом. Кроме того, целом брахиопод окружает кишку и заходит в прикрепительный стебелек.
У разных животных целом образуется по-разному (рис. 3). У вторичноротых — иглокожих, полухордовых и хордовых — стенка первичной кишки образует карманообразные выпячивания, которые затем отделяются от нее и становятся замкнутыми мешками. Полость внутри этих мешков и есть целом. Такой способ образования целома называется энтероцельным (от греческого энтерон — кишка). В этом случае целомическая полость происходит прямо от кишечной полости. Но у первичноротых целом, как правило, образуется по-другому. Сначала по бокам от кишки обособляются компактные группы клеток, не имеющие никаких полостей. Только потом эти клетки расходятся, и внутри каждой их кучки появляется полость, которая становится целомом. Такой способ образования целома называется шизоцельным (или схизоцельным).
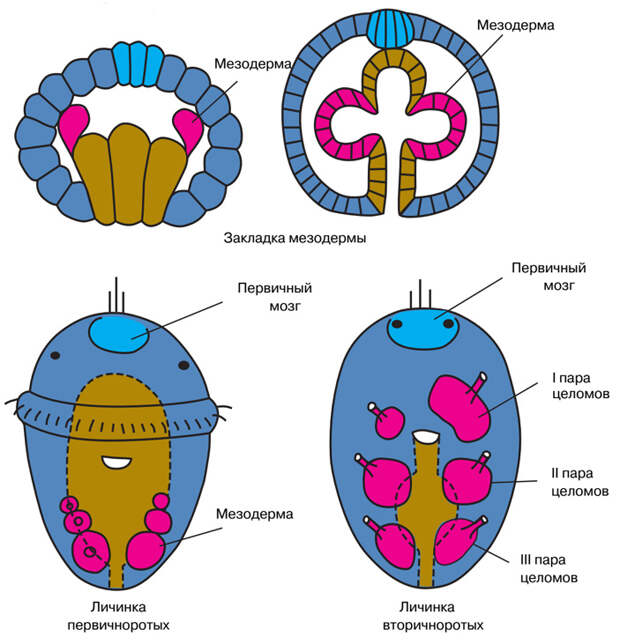
Почему это важно? Карманы первичной кишки, из которых развивается целом у вторичноротых, очень напоминают разделенные перегородками (септами) карманы кишечника коралловых полипов. Напрашивается предположение, что именно из таких карманов и образовались целомические мешки (см. рис. 4 в статье Проблема гомологии конечностей: от щупалец кораллов до крыльев насекомых, «Элементы», 23.01.2023). Тогда энтероцельный способ развития целома можно считать кратким повторением (рекапитуляцией) его эволюционного возникновения. Так родилась энтероцельная теория происхождения целома, которая, в свою очередь, очень важна для проблемы происхождения двусторонне-симметричных животных.
Чаще всего энтероцельную теорию связывают с именами братьев Оскара и Рихарда Гертвигов (Oscar Hertwig, Richard Hertwig), которые в 1881 году выпустили целую книгу «Теория целома» (O. Hertwig, R. Hertwig, 1881. Die Coelomtheorie). Но приоритет, видимо, принадлежит британскому биологу Рэю Ланкестеру (Edwin Ray Lankester), тому самому, который был одним из прототипов конан-дойловского профессора Челленджера (E. R. Lankester, 1877. Notes on embryology and classification for the use of students). Защитником энтероцельной теории был немецкий зоолог Адольф Ремане (Adolf Remane), который суммировал аргументы за и против нее в обстоятельной статье (A. Remane, 1963. The enterocelic origin of the celom). Давайте посмотрим, как эти аргументы выглядят сейчас.
Теория целома
Как мы уже знаем, развивающиеся целомические мешки некоторых двусторонне-симметричных животных сильно напоминают карманы кишечника коралловых полипов. Но при чем тут коралловые полипы? Если они близки к предкам двусторонне-симметричных животных, то должны быть самой древней и примитивной (базальной) группой стрекающих. А это вызывало большие сомнения. Например, американская исследовательница Либби Хайман (Libbie Henrietta Hyman), прославившаяся тем, что написала в одиночку превосходное шеститомное руководство по зоологии беспозвоночных, считала, что базальная группа стрекающих — это гидроиды, потому что они устроены гораздо проще, чем кораллы. Но этот аргумент Хайман учитывает только строение стадии полипа. Между тем у всех классов стрекающих, кроме кораллов, есть еще и подвижная стадия медузы, которая всегда устроена более-менее сложно. Если бы коралловые полипы произошли от гидроидов, они должны были бы вторично потерять стадию медузы, а такое предположение ничем не подтверждается и не добавляет гипотезе Хайман убедительности.
Кроме того, простота далеко не всегда означает примитивность. Здесь Ремане приводит изящный ботанический пример. Трудно представить более простое цветковое растение, чем ряска. Но это вовсе не свидетельствует о примитивности: ряска — чрезвычайно сильно специализированный представитель ароидных, которые, как правило, имеют нормальные стебли и листья, да и само семейство ароидных занимает среди цветковых далеко не базальное положение. В зоологии тоже много подобных случаев, и простота гидроидных полипов вполне может оказаться вторичной.
Спор между Ремане и Хайман разрешила молекулярная филогенетика. В 1992 году выяснилось, что коралловые полипы — единственный класс стрекающих, у которого митохондриальная ДНК остается кольцевой (D. Bridge et al., 1992. Class-level relationships in the phylum Cnidaria: evidence from mitochondrial genome structure). У всех остальных стрекающих митохондриальная ДНК линейная. Митохондрии, как известно, произошли от альфа-протеобактерий, некогда вселившихся в клетку архейного предка эукариот. У бактерий ДНК кольцевая. У большинства животных она осталась кольцевой и в митохондриях. Переход митохондриальной ДНК в линейную форму — нечастое эволюционное новшество. Тот факт, что у коралловых полипов митохондриальная ДНК осталась кольцевой, свидетельствует, что они и есть базальная группа стрекающих. Дальнейшие молекулярные исследования подтвердили этот вывод (см. Двусторонняя симметрия кораллов древнее радиальной, «Элементы», 26.10.2016). Линеаризация митохондриальной ДНК и появление стадии медузы — особенности продвинутых стрекающих, которые не были свойственны первым членам этой группы. Общий предок стрекающих и двусторонне-симметричных животных (билатерий) вполне мог быть похож на кораллового полипа.
Свое слово сказала и эволюционная биология развития. Выяснилось, например, что ген Brachyury, который у билатерий, в частности, регулирует развитие осевой мезодермы (см. У одноклеточных организмов есть ген, способный управлять развитием хорды, «Элементы», 25.10.2013), у коралловых полипов экспрессируется в септах, разделяющих кишечные карманы, то есть именно там, где ему и следовало бы экспрессироваться согласно энтероцельной теории (C. Scholtz, U. Technau, 2003. The ancestral role of Brachyury: expression of NemBra1 in the basal cnidarian Nematostella vectensis (Anthozoa)). Есть и другие косвенные генетические подтверждения того, что кишечные карманы кораллов и целомические мешки билатерий могут быть гомологами (см. Проблема гомологии конечностей: от щупалец кораллов до крыльев насекомых, «Элементы», 23.01.2023).
Другое возражение против энтероцельной теории имеет более общий характер. Дело в том, что в процессах формообразования (морфогенеза) животных организмов есть два главных участника: эпителий и мезенхима. Эпителий — это плотный слой клеток, выстилающий некоторую поверхность. Клетки эпителия сидят на общей базальной мембране и связаны между собой межклеточными контактами. Двигаться независимо друг от друга они не могут. Мезенхима — это рыхлая соединительная ткань, в которой клетки могут более-менее свободно двигаться в разные стороны. Эпителий хорош тем, что может упорядоченно менять свою форму: впячиваться, выпячиваться, образовывать полости, трубки и более сложные структуры. Клетки мезенхимы перемещаются по организму гораздо более хаотично. Кроме того, эпителий и мезенхима способны превращаться друг в друга. Эпителий может рассыпаться в мезенхиму, но и мезенхима может самоорганизоваться в эпителий.
Вклад эпителия и мезенхимы в формирование одних и тех же органов может сильно различаться у разных животных. Развитие целома — яркий тому пример. При энтероцельном способе развития целома целомический эпителий непосредственно «отпочковывается» от кишечного эпителия, а при других способах целомический эпителий самоорганизуется из мезенхимы. Какой из этих вариантов эволюционно первичен? Очень похожие вопросы возникают и при исследовании развития других органов (например, первичной кишки и нервной трубки).
Мнения зоологов по этим вопросам резко разошлись. Например, Владимир Николаевич Беклемишев считал эпителиальный морфогенез более организованным, а следовательно — возникшим позже. «Эпителий легче проделывает все движения, ведущие к закладке органов... как военный отряд маневрирует легче толпы» (В. Н. Беклемишев, 1925. Морфологическая проблема животных структур). Гертвиги и Ремане, наоборот, считали, что у древнейших многоклеточных животных всё, что только можно, происходило путем выгибания, прогибания или отпочковывания эпителиальных пластов. Ремане резонно указал, что мезенхимный морфогенез на самом деле сложнее эпителиального. В нём больше этапов: клетки мезенхимы должны выселиться из исходного эпителия, мигрировать россыпью к месту назначения и там заново самоорганизоваться в эпителий. Поэтому следовало бы ожидать, что эпителиальный морфогенез окажется более примитивным.
На сегодняшний день можно утверждать: в тех случаях, когда возможна проверка, факты оказываются скорее на стороне Гертвигов и Ремане. Например, среди вторичноротых больше всего примитивных признаков сохраняют кишечнодышащие, у которых формообразование эпителизовано максимально сильно: животное буквально представляет собой «сверток эпителиев» (В. В. Малахов, О. В. Ежова, 2013. О саккоглоссусе, зоологах и перевернутых хордовых). Среди стрекающих, как мы знаем, базальное положение занимают кораллы, у которых формообразование очень сильно эпителизовано по сравнению с теми же гидроидами. Энтероцельная теория происхождения целома хорошо вписывается в эту логику.
Целом брахиопод
Как образуется целом у брахиопод?
В классической зоологии было принято считать, что у брахиопод, в отличие от подавляющего большинства других первичноротых, целом образуется энтероцельным путем, то есть из карманов первичной кишки. Этот момент подчеркивали и Ланкестер, и Ремане; оба они рассматривали энтероцельное образование целома как очень древний признак, унаследованный от первых билатерий (см. ниже, рис. 5). Однако в 2000 году вышла работа, в которой на современном уровне, с использованием электронного микроскопа, было показано, что у двух изученных видов брахиопод целом образуется не из кишечной полости, а благодаря расхождению клеток исходно плотных мезодермальных зачатков, то есть шизоцельно (C. Lüter, 2000. The origin of the coelom in Brachiopoda and its phylogenetic significance). Естественно, это поставило старые представления под угрозу.
Чтобы выяснить истину, российские исследовательницы Татьяна Валерьевна Кузьмина и Елена Николаевна Темерева изучили развитие брахиоподы Coptothyris grayi, живущей в Японском море. Они выводили личинок брахиоподы в лаборатории и детально исследовали их строение на разных стадиях с помощью с помощью конфокального лазерного сканирующего микроскопа (рис. 4).

Личинка Coptothyris grayi выходит из яйца на стадии поздней бластулы, имеющей шарообразную форму. Переход к стадии гаструлы происходит путем инвагинации, то есть впячивания одного из полушарий зародыша, которое, погружаясь внутрь, образует первичную кишку (рис. 4a). Вскоре после того, как симметрия зародыша превращается из радиальной в двустороннюю, первичная кишка образует две пары выступающих карманов: переднюю и заднюю. Это и есть зачатки целомов. На этой стадии кишечная полость непосредственно продолжается в полости целомических карманов (рис. 4b, 4c), а это означает, что способ формирования целома у Coptothyris grayi — энтероцельный.
На более поздней стадии личинка брахиоподы становится трехлопастной (рис. 4d). В ней выделяются головная лопасть, зачаток мантийной лопасти и педальная лопасть, из которой впоследствии развивается прикрепительный стебелек. При этом передний целомический карман становится целомом головной лопасти, а задний целомический карман делится на два целома, которые размещаются в мантийной и педальной лопастях (рис. 4e, 4f). Таким образом, целомическая мезодерма Coptothyris grayi происходит из двух источников — переднего и заднего.
Нужно добавить, что у брахиопод (во всяком случае, у тех, которые были объектами обсуждаемых исследований) личинки лецитотрофные: это означает, что они не питаются самостоятельно, а питаются запасенным в клетках желтком. Поэтому клетки у них большие, а свободные пространства между клетками — не очень. Это и может быть причиной того, что не у всех брахиопод удалось обнаружить зачаточные целомические полости.
Кроме того, вполне вероятно, что в процессе эволюции брахиопод у некоторых из них энтероцельный способ закладки целома постепенно превратился в шизоцельный. Такие эволюционные превращения хорошо документированы (см. ниже). О брахиоподах еще Хайман писала: «Итак, имеющиеся данные указывают, что у беззамковых брахиопод целом шизоцельный, а у замковых — энтероцельный» (L. H. Hyman, 1959. The Invertebrates. Volume V: Smaller coelomate groups). Сейчас эта классификация устарела, но Coptothyris grayi по ней относится как раз к замковым брахиоподам.
Два источника мезодермы
Кузьминой и Темеревой удалось показать, что мезодерма у Coptothyris grayi формируется из двух независимых источников — переднего и заднего. Для брахиопод это показано впервые. Зато два источника эмбриональной мезодермы были раньше описаны у многих других животных, например у плоских червей, немертин, моллюсков, ракообразных и форонид. В работах, посвященных форонидам, Елена Николаевна Темерева и Владимир Васильевич Малахов предполагают, что «передний и задний источники мезодермы по своему положению связаны с ротовым и анальным отверстиями, которые согласно построениям сравнительных анатомов представляют собой продукты разделения щелевидного бластопора радиально-симметричного предка» (Е. Н. Темерева, В. В. Малахов, 2007. Развитие и филогения форонид; см. также посвященную этой теме презентацию). Бластопор — это, согласно принятой здесь интерпретации, зародышевый первичный рот. Однако здесь не все так просто, и нам стоит сделать краткий обзор имеющих отношение к делу фактов.
Начнем с форонид. У этих щупальцевых животных, родственников брахиопод, личинка имеет три отдела целома: предротовой (протоцель), щупальцевый (мезоцель) и туловищный (метацель) (Е. Н. Темерева, В. В. Малахов, 2006. Ответ Томасу Бартоломеусу: Личинка форониды Phoronopsis harmeri Pixell, 1912 имеет тримерную организацию целома). В терминах классической зоологии такой план строения называется архицеломатным или архимерным (рис. 5). Многие зоологи — например, тот же Ремане — считали, что архимерное строение является исходным для всех билатерий и что форониды могут служить его образцом. Это предположение служит основой так называемой архицеломатной гипотезы происхождения билатерий (см. В. В. Малахов, 2004. Новый взгляд на происхождение билатерий). Мезодерма у изученных видов форонид происходит из двух зачатков — переднего и заднего. Передняя мезодерма возникает из передней стенки первичной кишки, которую она подковообразно огибает, а задняя отделяется от кишки ближе к анальному отверстию. Из переднего мезодермального зачатка образуется выстилка предротового и щупальцевого целомов, а из заднего — выстилка туловищного целома. Необходимо добавить, что у некоторых форонид предротовой целом (протоцель) редуцируется и целомов остается только два: щупальцевый и туловищный.
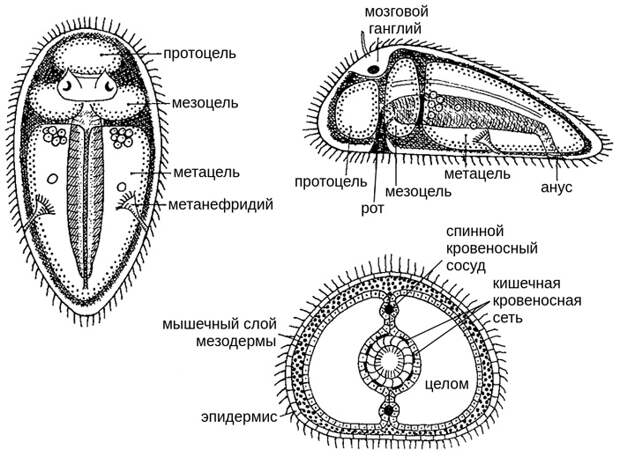
У брахиопод предротовой целом (протоцель) редуцирован очень сильно. Обнаружить его пока удалось только у беззамковой брахиоподы Lingula (E. N. Temereva et al., 2015. Demonstration of the preoral coelom in the brachiopod Lingula anatina with consideration of its phylogenetic significance). У подавляющего большинства брахиопод передний целомический зачаток дает начало только целому лофофора. Таким образом, у Coptothyris grayi головной целом — это мезоцель, а мантийный и педальный целомы — части разделившегося метацеля. Три целома личинки брахиоподы — это не те три целома, которые были у архимерных животных (хотя преемственность от них проследить можно). Так же, как и у форонид, мезоцель у брахиопод развивается из переднего мезодермального зачатка, а метацель — из заднего. Что касается разделения метацеля перегородками на дополнительные целомы, то у щупальцевых животных это не редкость. Остатки этих перегородок есть у взрослых брахиопод, они называются латеральными мезентериями (см. рис. 2, на котором метацель подразделен даже на три целома, и статью E. N. Temereva, V. V. Malakhov, 2011. The evidence of metamery in adult brachiopods and phoronids). Подразделение метацеля можно рассматривать как начальный этап формирования сегментации, примерно такой, как сегментация кольчатых червей; иное дело, что у щупальцевых этот процесс далеко не зашел.
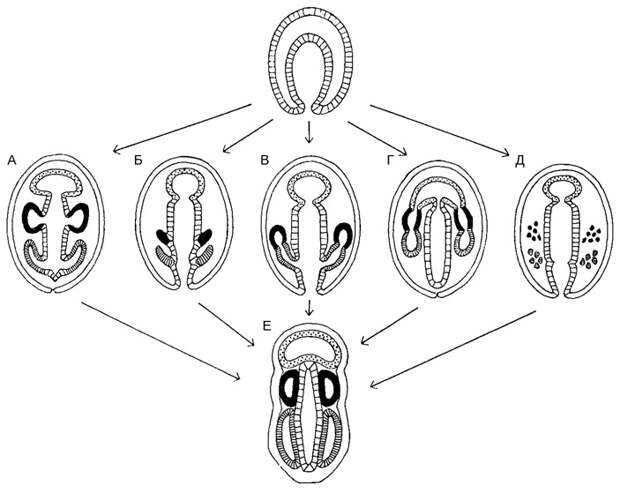
Еще одна группа животных, которой свойственно архимерное строение, — полухордовые, в частности уже упоминавшиеся кишечнодышащие. Это образцовые архицеломатные животные. Область протоцеля у них называется хоботком, область мезоцеля воротничком, а область метацеля туловищем. Никогда не подвергалось сомнению, что для кишечнодышащих в общем характерно энтероцельное образование целомов. Но, как известно, дьявол живет в деталях, а детали здесь на удивление разнообразны (рис. 6; О. М. Иванова-Казас, 1978. Сравнительная эмбриология беспозвоночных животных: Иглокожие и полухордовые). Есть виды кишечнодышащих, у которых целомы хоботка, воротничка и туловища образуются идеально правильным способом — как независимые друг от друга полые выпячивания стенки первичной кишки (рис. 6А). Есть виды, у которых зачатки целомов сначала обособляются от первичной кишки в виде плотных разрастаний, а уже потом в них возникают полости (рис. 6Б). В этом случае энтероцельный способ образования целома сменяется на шизоцельный. Есть виды, у которых целомы образуются из двух зачатков: переднего, дающего только хоботковый целом, и заднего, который делится на воротничковый и туловищный целомы (рис. 6В). Это, как видим, напоминает развитие целомов форонид и брахиопод. Есть виды, у которых целомы воротничка и туловища отрастают от уже обособившегося целома хоботка (рис. 6Г). В этом случае источник целомической мезодермы всего один. Наконец, есть виды, у которых целомы самоорганизуются из мезенхимы (рис. 6Д). Как видим, и энтероцельность/шизоцельность, и число и расположение источников целомической мезодермы — это признаки, которые могут довольно легко меняться в ходе эволюции, причем не влияя на конечный результат морфогенеза: «готовые» целомы у всех кишечнодышащих устроены примерно одинаково (рис. 6Е).
Еще один пример, который приводят Кузьмина и Темерева (и академик Малахов в других работах), относится к позвоночным животным. В самой передней части тела у зародышей позвоночных находится прехордальная пластинка, которая подстилает передний и промежуточный мозг и которую можно рассматривать как особый источник мезодермы. При желании это позволяет распространить теорию «двух источников» на позвоночных животных. Но и тут не все так просто. Прежде всего надо учитывать, что архимерное подразделение тела у позвоночных сохранилось, хотя и в замаскированном виде. Позвоночные — сегментированные, или, правильнее говоря, метамерные животные: это проявляется у них в строении осевого скелета, жаберного аппарата, мышц, нервов и некоторых других органов. Области протоцеля у позвоночных соответствует предчелюстной метамер, области мезоцеля — челюстной метамер, а области метацеля — все остальное тело (см. Изменчивость змеиного черепа документирует его эволюционные перестройки, «Элементы», 03.06.2020). При этом все метамерные мезодермальные образования у позвоночных развиваются из осевой (параксиальной) мезодермы, в состав которой не входит только зачаток хорды: он непарный, расположен строго посредине, и метамерия его не затрагивает. Что же касается прехордальной пластинки, то она, во-первых, находится в зоне предчелюстного метамера, и во-вторых, является непосредственным продолжением того самого зачатка хорды (рис. 7; H. C. Bjerring, 1971. The nerve supply to the second metamere basicranial muscle in osteolepiform vertebrates, with some remarks on the basic composition of the endocranium). В эмбриологических экспериментах изолированную прехордальную пластинку тритона удавалось даже заставить дифференцироваться в ткань хорды (H. Takaya, 1953. On the notochord-forming potency of the prechordal plate in Triturus gastrulae). Эта потенция в ней сохранилась. В норме из прехордальной пластинки, по-видимому, развиваются некоторые зародышевые хрящи мозговой коробки. А вот к целомическим структурам она не имеет отношения.
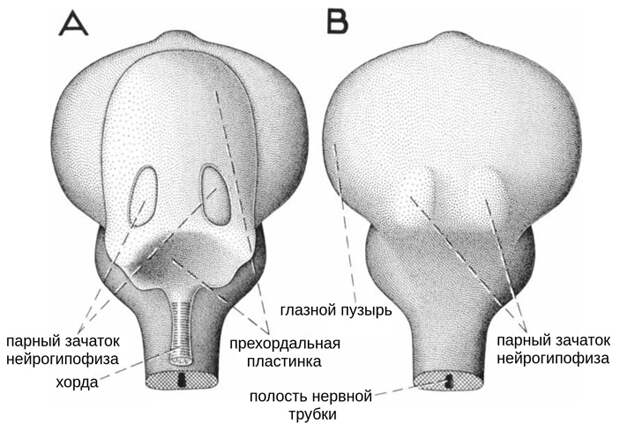
К этому можно добавить, что у самого базального современного хордового — ланцетника — прехордальной пластинки в принципе не может быть: соответствующая часть мезодермы у него становится продолжением хорды, которая (в отличие от хорды позвоночных) доходит до самого переднего конца тела. Таким образом, «два источника мезодермы» у позвоночных — это скорее аналогия, чем знак преемственности с тем, что наблюдается у брахиопод и форонид. Но и аналогия может свидетельствовать о том, что за планами строения разных животных скрываются глубокие общие закономерности.
Источник: T. V. Kuzmina, E. N. Temereva. Two sources of mesoderm in brachiopods // Doklady Biological Sciences. 2024. DOI: 10.1134/S0012496623700837.
Сергей Ястребов
Свежие комментарии