
Исследования биологии развития рыб позволили выдвинуть новую версию происхождения парных — грудных и брюшных — плавников, от которых, в свою очередь, произошли конечности наземных позвоночных. Предполагается, что предшественником парных плавников была непарная преанальная плавниковая складка (ПАПС), которая часто присутствует у личинок современных рыб. Данные по экспрессии регуляторных генов и по эмбриональному происхождению клеток, формирующих скелет, подтверждают, что ПАПС вполне может быть гомологична парным плавникам. Получается, что в результате удвоения ПАПС сначала возникли длинные брюшные плавники (как у ископаемых бесчелюстных группы анаспид), а потом от них «отпочковались» грудные.
Что такое плавники
Тело большинства позвоночных животных имеет подвижные придатки, которые называются конечностями. У наземных позвоночных конечности бывают только парными: это передние и задние лапы со всеми их видоизменениями (например, у птиц передние лапы превращены в крылья). У рыб передним лапам соответствуют грудные плавники, а задним лапам — брюшные плавники (рис. 2).
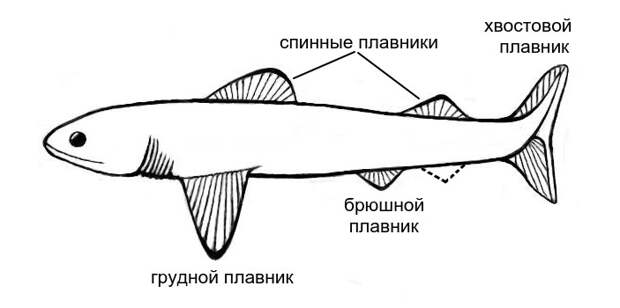
Кроме того, у подавляющего большинства рыб есть непарные плавники: один или два (редко больше) спинных и анальный. Последний расположен на средней линии брюшной стороны тела позади анального отверстия или отверстия клоаки. Наконец, у водных позвоночных есть хвостовой плавник, который продолжает собой туловище и обычно служит главным источником движения. Вполне вероятно, что все непарные плавники когда-то образовывали непрерывную складку, тянувшуюся вдоль спины, вокруг хвоста и вперед до анальной области.
Сейчас что-то подобное можно увидеть, например, у головастика.Сравнительные анатомы традиционно интересовались в основном парными плавниками позвоночных. Это понятно: парные плавники устроены сложнее, а их эволюционная история необыкновенно богата.
Элементы костного или хрящевого скелета парных плавников приняло делить на базальные (они же просто базалии) и радиальные (они же просто радиалии). Базальные элементы крупнее и находятся ближе к основанию плавника, радиальные отходят от них. Базалии и радиалии образуют внутренний скелет плавника, имеющий собственную мускульную «оснастку». Кроме того, у любого рыбьего плавника есть еще и внешний скелет. Его образуют кожные лучи, которые либо окостеневают, либо (у хрящевых рыб) на всю жизнь остаются мягкими, но прочными белковыми нитями. Плавник, полностью лишенный «оторочки» из кожных лучей, — это уже не плавник, а ласт. Похоже, что именно исчезновение кожных лучей было решающим шагом на пути от рыбьего плавника к наземной конечности (см. Новооткрытые рыбьи гены помогли понять, почему первые четвероногие были многопалыми, «Элементы», 09.07.2010). С точки зрения эволюции самих плавников, однако, наибольший интерес представляет как раз их внутренний скелет. Он очень многообразен и подкидывает ученым головоломные задачи.
В некоторых плавниках базальные элементы образуют одну-единственную членистую ось. Радиальные элементы отходят от этой оси или с двух сторон, или только с одной. В первом случае плавник получается листовидным, его внутренний скелет в целом симметричен. Такой плавник называется бисериальным архиптеригием (рис. 3а). Во втором случае плавник асимметричен, зато радиалии, находящиеся только с одной стороны, становятся гораздо более мощными и толстыми. Такой плавник называется унисериальным архиптеригием (рис. 3б). В современном мире по типу бисериального архиптеригия устроены плавники двоякодышащих рыб, а по типу унисериального — плавники латимерии.
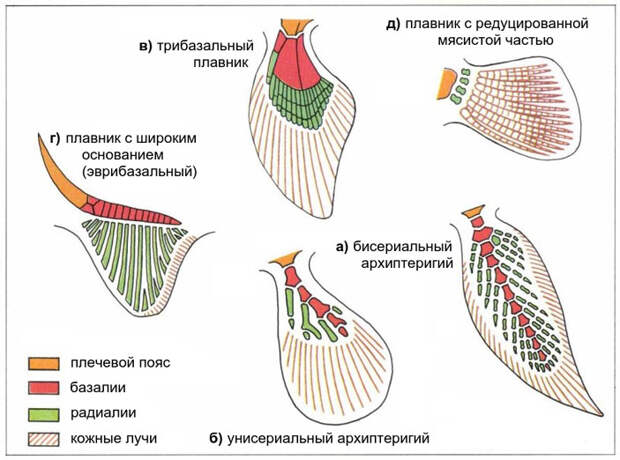
В учебниках обычно пишут, что конечность наземных позвоночных возникла из унисериального архиптеригия, но некоторые ученые считают, что из бисериального (см. D. N. Mednikov, 2014. Urodelans, Ichthyostega and the origin of the tetrapod limb). В качестве примера древней рыбы с бисериальным архиптеригием можно привести поролеписа (см. картинку дня Поролепис).
Другие плавники имеют в основании три параллельных (точнее, слегка веерообразно расходящихся) базальных элемента, от концов которых отходят более многочисленные радиалии. Такой плавник называется трибазальным (рис. 3в), и он характерен для современных акул. Кроме того, похожую конструкцию можно увидеть у одной архаичной группы костных рыб, а именно у многопёров.
Правда, плавники акул — не всегда строго трибазальные. В грудном плавнике у них иногда бывает и четыре базальных элемента. С другой стороны, в брюшном плавнике, который, как правило, устроен проще, базальных элементов обычно два, а то и всего один.
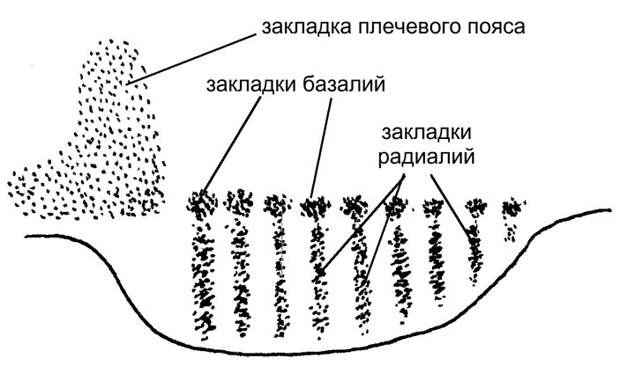
В эмбриональном развитии плавника акулы базалии образуются в результате слияния оснований радиалий (рис. 4). На ранних стадиях плавник устроен очень просто: он представляет собой широкую складку, в которую входит ряд параллельных радиалий. У взрослых современных рыб такой тип плавника в чистом виде не встречается. Но вот у ископаемой хрящевой рыбы Cladoselache, жившей примерно 360 миллионов лет назад, грудной плавник довольно близок к тому, что наблюдается у зародышей современных акул (рис. 3г, а также рис. 1). Он выглядит как широкий выступ, в который, по последним данным, заходят 13 тонких радиалий (T. Tomita, 2015. Pectoral fin of the Paleozoic shark, Cladoselache: new reconstruction based on a near-complete specimen). Еще в XIX веке была высказана и стала популярной идея, что грудной плавник Cladoselache воплощает древнейший тип плавников, общий для предков всех рыб.
У костистых рыб (96% современных видов рыб относится к этой группе) базальные и радиальные элементы плавников стали рудиментарными либо даже исчезли вовсе. Почти весь скелет такого плавника образован кожными лучами (рис. 3д). Это — эволюционно молодой тип плавника, облегченный за счет редукции внутреннего скелета и соответствующих мышц. В нем почти нет мясистой основы. Именно таковы полупрозрачные плавники, которые мы привыкли видеть у всевозможных речных и аквариумных рыб.
Конечность наземных позвоночных, конечно, могла произойти только от мясистого плавника — одного из архиптеригиев. Но откуда взялись сами парные плавники? Над этим вопросом сравнительные анатомы размышляют уже 150 лет, и он до сих пор не решен.
Теория Гегенбаура
Принципиальное отличие конечностей позвоночных от конечностей беспозвоночных (параподий кольчатых червей, ножек онихофор, тихоходок и членистоногих) состоит в следующем. Конечность беспозвоночного принадлежит одному сегменту тела. Позвоночные — тоже сегментированные (метамерные) животные, их сегментация проявляется в строении зародышевых закладок и многих внутренних органов — позвоночника, мышечной системы, нервов. Однако конечность позвоночного, будь то плавник или лапа, всегда образуется за счет нескольких сегментов, которые посылают в зачаток конечности свои нервы и мускульные почки. Поэтому напрямую сопоставлять конечности позвоночных и беспозвоночных нельзя. Это принципиально разные структуры.
У современных бесчелюстных — миног и миксин — парные плавники полностью отсутствуют, и нет никаких свидетельств, что у предков этих животных они когда-нибудь были. Поэтому парные конечности нельзя считать общим признаком позвоночных. Мы сейчас не учитываем многочисленные случаи, когда парные конечности вторично исчезли (у змей, безногих ящериц, червяг, мурен и некоторых ископаемых животных). У миног и миксин отсутствие парных плавников, скорее всего, первично.
По этим причинам большинство биологов считает, что первые позвоночные не имели парных конечностей. Эти уникальные придатки появились только в ходе их дальнейшей эволюции. Но откуда же они все-таки взялись?
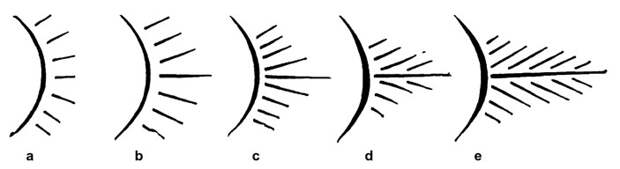
Первую серьезную гипотезу на этот счет высказал немецкий сравнительный анатом Карл Гегенбаур (Karl Gegenbaur; см. K. Gegenbaur, 1878. Elements Of Comparative Anatomy). Ее можно назвать теорией жаберного происхождения плавников.
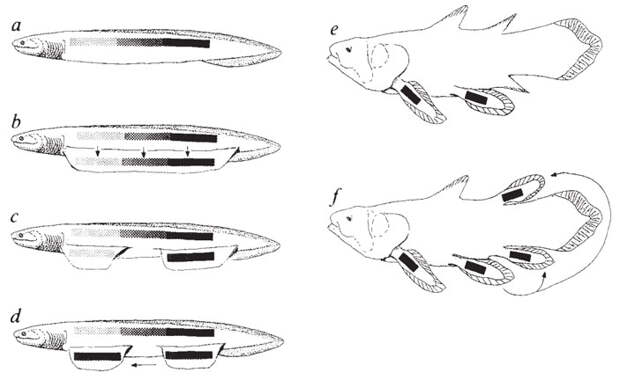
Дело в том, что на скелетных жаберных дугах у рыб есть выросты — жаберные лучи. Представим, что один луч получил особенно сильное развитие, и другие лучи стали вырастать уже на нем, а не на дуге (рис. 5). Если главный луч еще и сегментируется, разбившись на отдельные членики, то получится структура, очень похожая на бисериальный архиптеригий — плавник, присущий двоякодышащим рыбам. Именно этот тип плавника Гегенбаур считал исходным для позвоночных. Сама жаберная дуга при этом должна была превратиться в пояс конечности.
Гипотеза Гегенбаура не имеет никаких палеонтологических подтверждений. Ей нельзя отказать в красоте, но она основана на фактах, которые были известны автору в XIX веке, и на серии изящных домыслов. Дальнейшее развитие сравнительной анатомии и эмбриологии заставило выдвинуть против этой гипотезы сильные доводы. Например, сейчас хорошо известно, что пояса конечностей и жаберные дуги имеют совершенно разное эмбриональное происхождение: плечевой и тазовый пояса вместе со скелетом конечностей развиваются из мезодермы, а скелет жаберных дуг — из нервного гребня (см., например: «Четвертый зародышевый листок» позвоночных зародился у низших хордовых, «Элементы», 04.02.2015). К тому же теория Гегенбаура никак не объясняет важнейшую особенность конечностей позвоночных — то, что каждая из них развивается из материала нескольких сегментов тела. Конечность, образовавшаяся по Гегенбауру, должна была бы принадлежать только одному сегменту — тому, к которому относилась исходная жаберная дуга. Реальные конечности позвоночных никогда не бывают так устроены.
Правда, в последнее время появились данные из области генетики развития, показывающие, что в идеях Гегенбаура все же есть кое-что полезное. Существуют гены, которые регулируют, с одной стороны, развитие конечностей наземных позвоночных, а с другой — развитие жаберных дуг и жаберных лучей у современных рыб, причем механизмы производимой этими генами регуляции на уровне клеточных масс выглядят очень похоже (J. A. Gillis, B. K. Hall, 2016. A shared role for sonic hedgehog signalling in patterning chondrichthyan gill arch appendages and tetrapod limbs). Это гены Shh (Sonic hedgehog) и FGF8, кодирующий один из факторов роста фибробластов. Зашла речь даже о глубокой гомологии между жаберными лучами и парными конечностями (J. A. Gillis et al., 2009. Shared developmental mechanisms pattern the vertebrate gill arch and paired fin skeletons). Но это вряд ли означает, что плавники действительно произошли от жаберных лучей. Скорее всего, дело было по-другому. Гены Shh и FGF — это универсальные переключатели, способные принимать участие в самых разных процессах эмбрионального развития (см., например: Развивающиеся части спинного мозга сначала регулируются централизованно, а затем переходят на самоуправление, «Элементы», 01.10.2014). Жаберный аппарат эволюционно возник раньше, чем парные конечности. Нет ничего удивительного в том, что модель регуляции развития скелетных структур генами Shh и FGF8, оформившись в жаберном аппарате, потом была с еще большим успехом использована в новых органах — парных плавниках.
Теория боковой складки
На протяжении ста с лишним лет самой популярной версией происхождения парных плавников была теория боковой складки, которую предложил в XIX веке Фрэнсис Бальфур (Francis Maitland Balfour).
Исследуя развитие акулы, Бальфур заметил, что у ее зародыша между зачатками грудных и брюшных плавников проходит полоса эпителия c характерно вытянутыми столбчатыми клетками, очень похожая на продолжение плавника. Он предположил, что это остаток единой складки, впоследствии распавшейся. Плавниковые лучи, изначально многочисленные и примерно одинаковые (как у Cladoselache), в конце концов срослись в мощные базальные элементы основания плавника. Так и возник трибазальный плавник современных акул, который, по Бальфуру, гораздо древнее всевозможных архиптеригиев.
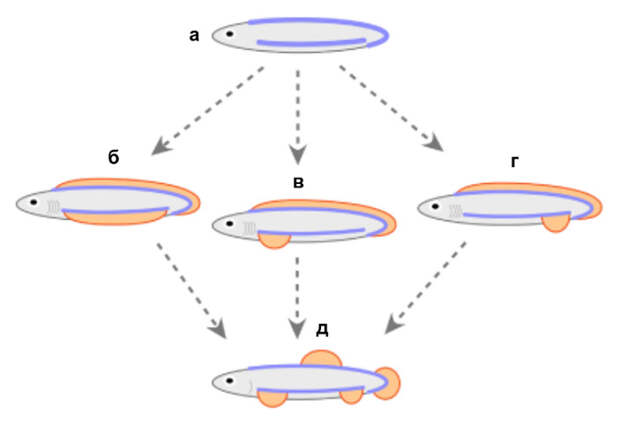
Тут вырисовывается следующий эволюционный сценарий (рис. 6а–б). Первоначально у позвоночных были только непарные плавники — скорее всего, в виде сплошной складки (см., например, изображение хайкоуихтиса). Но без дополнительных стабилизаторов движение такого животного было плохо управляемо, примерно как движение современного головастика. Поэтому по бокам туловища появились горизонтальные стабилизирующие кили, которые помогали избежать нежелательного вращения тела вокруг продольной оси — проще говоря, заваливания то на один, то на другой бок. Это и были плавниковые складки. Но и такая конструкция оказалась не слишком удобной: при изгибах тела, которыми всегда сопровождается плавание водных позвоночных, складка с одной стороны растягивались, а с другой собиралась гармошкой. Поэтому сплошные складки распались на отдельные плавники, из которых в конце концов сохранились только самые передние и самые задние — грудные и брюшные.

Данные генетики развития и здесь расцвечивают старую гипотезу новыми красками. У позвоночных есть семейство регуляторных генов, которое называется Tbx. Оно относится к древней группе T-боксных генов (см. У одноклеточных организмов есть ген, способный управлять развитием хорды, «Элементы», 25.10.2013). У зародыша акулы в грудном плавнике экспрессируется ген Tbx5, а в брюшном — ген Tbx4 (M. Tanaka et al., 2002. Fin development in a cartilaginous fish and the origin of vertebrate limbs). Легко предположить, что некогда в древности существовал единый ген Tbx4/5, который экспрессировался длинной полосой вдоль всего бока. Потом он дуплицировался, и после распада боковой складки функции двух генов разделились (рис. 6е–з). И действительно, у ланцетника есть ген Tbx4/5, который экспрессируется полосой вдоль боковой мезодермы. Экспериментально показано, что белковый продукт этого гена может у зародыша позвоночного заменить продукты как гена Tbx4, так и гена Tbx5 (см. C. Minguillon et al., 2009. Tbx4/5 gene duplication and the origin of vertebrate paired appendages). Анализ нуклеотидных последовательностей подтверждает, что гены Tbx4 и Tbx5 возникли в результате дупликации гена Tbx4/5. Очевидно, это произошло при одной из полногеномных дупликаций, которыми было отмечено начало эволюции позвоночных (см. А. В. Байрамов и др., 2021. Геномные дупликации как основа эволюционного успеха позвоночных).
Теория боковой складки выглядит убедительно и давно вошла в учебники. Некоторые авторы даже считали, что это не теория, а твердо установленный факт. Но надо учитывать, что абсолютно все доводы в пользу этой теории — косвенные (см., например: В. Г. Борхвардт, 2016, первая публикация в 1986. Происхождение парных плавников: состояние проблемы). Никто никогда не видел позвоночного, у которого хотя бы в эмбриональном состоянии действительно была бы непрерывная боковая плавниковая складка. Ископаемых находок такого рода тоже нет. В этом смысле вопрос остается открытым.
Теория плавниковых шипов
Американский палеонтолог Уильям Кинг Грегори (William King Gregory) считал обязательным элементом первых парных плавников шипы. Почему шипы? Дело в том, что это чрезвычайно характерный элемент плавников древних (палеозойских) позвоночных. Есть целая группа вымерших рыб, которая обязана шипам своим названием, — акантоды (от греческого ακανθα — шип, колючка). Акантоды не относятся ни к хрящевым, ни к костным рыбам, а представляют собой особую группу, очень примитивную по некоторым скелетным признакам. Все плавники акантод, кроме хвостового — два спинных, грудной, брюшной и анальный — имеют на своем переднем краю массивные шипы из твердого материала. Очень похожие шипы встречаются у панцирных рыб (еще одна полностью вымершая большая группа), у ископаемых акул, у некоторых бесчелюстных — словом, во всех древнейших группах позвоночных. Гипотезу Грегори можно назвать теорией плавниковых шипов. Она неплохо соответствует общей тенденции эволюции ранних позвоночных, известной из палеонтологической летописи: сначала у них активно развивался кожный скелет, образовывавший всевозможные шипы и панцири, а потом кожный скелет стал редуцироваться и замещаться внутренним.
Следует отметить, что Грегори вовсе не думал, будто первой стадией эволюции плавника был голый шип. Типичный плавник акантоды состоит из двух компонентов: впереди — твердый шип, а за ним — мясистая лопасть с хрящевым скелетом, слегка напоминающим скелет трибазального плавника акулы. Именно такой тип плавника и был исходным по мнению Грегори (W. K. Gregory, H. C. Raven, 1941. Studies on the origin and early evolutin of paired fins and limbs). В дальнейшем шип исчез, мясистая часть плавника разрослась, и получились «нормальные» рыбьи плавники, существующие до наших дней (рис. 7).
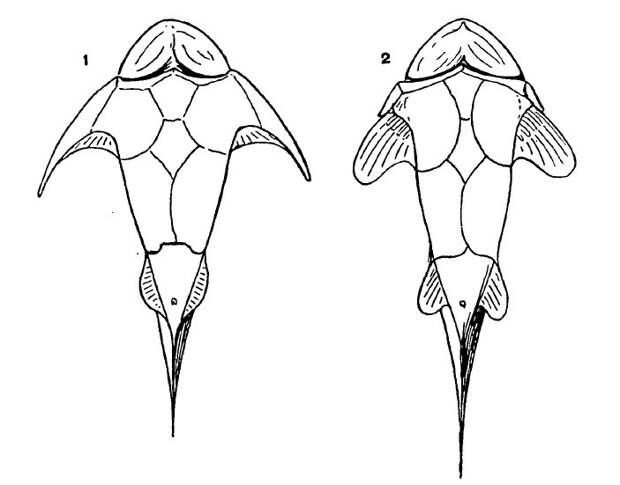
Признак акантод, не встречающийся ни у каких других позвоночных — множественность парных плавников. У многих (хотя и не у всех) родов акантод между грудными и брюшными плавниками находится до пяти пар дополнительных небольших плавничков, тоже снабженных шипами (см. рис. 12). Таким образом, парных конечностей у них несколько пар. Этот факт часто приводят в подтверждение теории боковой складки — ведь дополнительные плавнички легко проинтерпретировать как ее остатки. Но тут не все так просто: некоторые палеонтологи считают, что образование дополнительных парных плавников у акантод — скорее всего, вторичное явление (см. Л. П. Татаринов, 1987. Очерки по теории эволюции).
Грегори не отрицал теорию боковой складки, а считал свою гипотезу в принципе совместимой с ней. Тем не менее теория боковой складки (в ее классической версии) и теория Грегори в некоторых случаях приводят к разным выводам. Плавники девонской хрящевой рыбы Cladoselache, имеющие очень широкое основание (см. рис. 2), по классической теории боковой складки примитивны, а по теории Грегори — нет. Состояние с большим числом хрящевых лучей и их распространением на весь широкий плавник Грегори трактует как специализацию. По его мнению, плавники Cladoselache не более примитивны, чем, например, грудной плавник ската — донной хрящевой рыбы с плоским и широким телом, у которой грудные плавники колоссально разрослись.
Конечно, плавники с шипами были относительно малоподвижны. Но и плавники с широкими основаниями тоже были малоподвижны. Это не влияло на их исходную функцию — служить горизонтальными стабилизирующими выростами. Тут шипы, укреплявшие плавники, были даже полезны.
Что первично?
Генетика развития, начавшая бурно развиваться в 1990-е годы, добавила к знаниям биологов о конечностях много нового.
В первую очередь новые данные касались генов семейства Hox. Эти универсальные регуляторные гены обычно контролируют у многоклеточных животных «разметку» отделов туловища (см., например: Развитие «сегментов» у книдарий контролируется Hox-генами, как у билатерий, «Элементы», 01.10.2018). Судьба частей тела, расположенных впереди или сзади, зависит от того, какие Hox-гены в них экспрессируются. Впереди экспрессируется меньше всего Hox-генов, сзади — больше всего. В хромосоме Hox-гены обычно собраны в единый кластер, и гены в этом кластере расположены удивительным образом в том же порядке, что и зоны их экспрессии в теле зародыша (принцип коллинеарности).
В зачатках конечностей позвоночных экспрессия Hox-генов тоже есть. Экспериментальная биология развития быстро выяснила, что в зачатках передних и задних конечностей экспрессируется один и тот же набор Hox-генов, и это гены «задней» группы, находящиеся ближе к концу кластера (Hox11–Hox13). Они же экспрессируются и в туловище на том уровне, на котором расположены задние конечности. А вот там, где растут передние конечности, в туловище активны совсем другие Hox-гены. «На своем месте» относительно туловища находятся задние конечности, но не передние (рис. 8).
Такие данные прямо-таки провоцировали предположение, что сначала у позвоночных появились только брюшные плавники. Потом они удвоились, и дополнительная пара плавников переместилась вперед, превратившись в грудные, но сохранив древний паттерн экспрессии Hox-генов (C. Tabin, E. Laufer, 1993. Hox genes and serial homology). Так появилась теория первичности брюшных плавников.
В противовес к ней была тут же предложена и теория первичности грудных плавников (M. Coates, 1993. Hox genes, fin folds and symmetry). Она основана на том, что у многих ископаемых бесчелюстных, в частности у членов группы Osteostraci (которых многие авторы считают ближайшими родственниками челюстноротых), есть только грудные плавники, причем превосходно развитые (см. изображение цефаласписа). Экспрессию в них «задних» Hox-генов можно было бы объяснить тем, что, когда парные плавники возникали, туловище было очень коротким: большая, плоская, закованная в панцирь голова древнего бесчелюстного почти сразу переходила в хвост.
А что же теория боковой складки? Ее сторонникам генетика развития дала серьезный повод для раздумий. Теория боковой складки предсказывает совершенно иную картину экспрессии Hox-генов, чем реально наблюдаемая. Действительно, если плавниковая складка первоначально тянулась вдоль всего туловища, то и в передних, и в задних конечностях должны экспрессироваться те же Hox-гены, что в туловище на соответствующем уровне. На самом деле все оказалось по-другому. Никакого общепринятого объяснения этот факт пока не имеет.
Правда, именно у позвоночных ситуация с Hox-генами сложнее, чем у большинства других животных. Дело в том, что в начале эволюции позвоночных происходили полногеномные дупликации (см. У мечехвостов обнаружены свидетельства полногеномной дупликации, «Элементы», 27.11.2015), поэтому Hox-кластеров у них несколько, например у наземных позвоночных — четыре. Они обозначаются буквами A, B, C и D. В зачатках конечностей экспрессируются Hox-гены кластеров A, C и D. Описанный тип экспрессии, когда и в передних, и в задних конечностях экспрессируются Hox-гены «задней» группы, наблюдается в кластерах A и D. А вот кластер C ведет себя в согласии с теорией боковой складки. Почему так — загадка.
Парное из непарного
Наконец, следует упомянуть теорию происхождения парных плавников от непарных. Она была высказана в XIX веке в работах викторианских анатомов Джона Гудсира (John Goodsir) и Джорджа Мюррея Хамфри (George Murray Humphry). Дело в том, что у животных очень мало истинно непарных органов. Почти любой непарный орган возникает в эмбриональном развитии из парных элементов и может превратиться в парный, если что-то помешает этим элементам слиться. Это относится и к непарным плавникам. Их скелет и особенно мускулатура формируются из парных зачатков, которые смыкаются на средней линии (см. Ф. Я. Дзержинский, 2005. Сравнительная анатомия позвоночных животных). По Хамфри, парные плавники можно рассматривать как гомологи непарного анального плавника, которым помешал слиться внутренностный мешок (G. M. Humphry, 1870. On the homological relations to one another of the mesial and lateral fins of the osseous fishes).
Эволюционная биология развития подтверждает, что парные и непарные плавники действительно имеют много общего. В развитии как парных, так и непарных плавников ключевую роль играет находящийся на вершине зачатка плавника апикальный эктодермальный гребень (apical ectodermal ridge, AER; см. Длинные плавники скатов – результат добавления новой точки роста, «Элементы», 22.12.2015). Развитие зачатка плавника и там и там регулируется примерно одними и теми же генами, в первую очередь — уже упоминавшимися генами семейства FGF (R. Freitas et al., 2006. Evidence that mechanisms of fin development evolved in the midline of early vertebrates).
Исследования по биологии развития показали, что у акулы все непарные плавники возникают сначала в виде единой непрерывной складки, идущей от спины до анального отверстия. Индивидуальное развитие, конечно, далеко не всегда повторяет историческое, но описания кембрийских хордовых (которых сейчас известно не так уж мало) подтверждают, что такое состояние, скорее всего, было для них исходным. Здесь напрашивалась гипотеза, которую высказала в 2010 году британская исследовательница Зерина Джохансон (Zerina Johanson). У первых позвоночных был сплошной непарный плавник, эмбриональное развитие которого определялось единственной срединной компетентной зоной (эмбриологический термин «компетенция» означает готовность ткани образовать определенный орган под действием подходящего стимула). Затем эта зона буквально дуплицировалась, породив пару аналогичных (или гомологичных?) компетентных зон по бокам тела. Там-то и стали закладываться парные плавники (Z. Johanson, 2010. Evolution of paired fins and the lateral somitic frontier).
Новые палеонтологические данные
Мы рассмотрели основные теории происхождения парных плавников. Теперь посмотрим, как влияют на выбор между этими теориями данные палеонтологии и биологии развития, полученные в начале XXI века.
Начнем с палеонтологии. В 2003 году была описана раннедевонская акула Doliodus, у которой грудной плавник имеет мощный шип и за ним — мясистую часть с несколькими хрящевыми лучами (рис. 9; R. F. Miller et al., 2003. The oldest articulated chondrichthyan from the Early Devonian period). На тот момент это была самая древняя хрящевая рыба, от которой удалось найти более-менее полный скелет (вернее, его отпечатки). Эта находка послужила основанием для предположения, что плавниковые шипы — общий признак всех челюстноротых. Очевидно, это прекрасно соответствует теории плавниковых шипов Грегори.

Детальное исследование плечевого пояса Doliodus показало, что его строение можно охарактеризовать как промежуточное между акулами и акантодами (J. G. Maisey et al., 2017. Pectoral morphology in Doliodus: bridging the "acanthodian’-chondrichthyan divide). Это тоже хороший довод за теорию Грегори. Ведь, как мы помним, именно акантоды прославились тем, что у них в состав плавников всегда входят шипы.
В 1998 году был открыт Psarolepis — рыба из самых низов девона, очень близкая к общему предку всех костных рыб (рис. 10; M. Zhu et al., 1999. A primitive fossil fish sheds light on the origin of bony fishes). Скелет его грудного плавника состоит из шипа и четырех хрящевых элементов. В дальнейшем выяснилось, что все примитивные костные рыбы имеют плавники с несколькими хрящевыми или костными лучами в основании и часто с шипами (M. Zhu, X. Yu, 2009. Stem sarcopterygians have primitive polybasal fin articulation). Это тоже хорошо соответствует теории Грегори.

В итоге можно реконструировать следующую последовательность событий. Плавник, исходный для всех рыб, имел твердый шип и примерно 4 луча внутреннего скелета. Именно таков плавник Psarolepis. При уменьшении числа лучей на один такой плавник становится трибазальным, как у раннедевонской костной рыбы Achoania, у многопёров и у современных акул. У большинства костных рыб шип исчезает, и дальнейшая эволюция идет только в мясистой части плавника. При редукции этой мясистой части получается плавник, почти целиком состоящий из кожной «оторочки» — как у костистых рыб. Прямо противоположный путь эволюции — исчезновение всех базальных лучей, кроме последнего, который разрастается и сегментируется. Так возникает плавник с членистой осью — унисериальный архиптеригий. В дальнейшем он легко (и, по-видимому, неоднократно) превращается в бисериальный.
Итак, похоже, что Грегори был прав. Последовательность эволюции плавников рыб, подтвержденная современной палеонтологией, оказалась ближе всего именно к его версии.
Но построения Грегори не решают проблему до конца. Во-первых, рыбы — это еще не все позвоночные. Кроме них есть еще и бесчелюстные. У современных бесчелюстных парных плавников нет, но у ископаемых — есть, и они бывают устроены очень необычно. Для полной картины это тоже надо учитывать.
Во-вторых, даже в отношении рыб теория Грегори неполна. Сравнительная анатомия делится на тектологию (она же органология) и архитектонику. Первая исследует отдельные части организма животного, вторая — план строения организма как целое. «Если тектология рассматривает особенности и типы повторяющихся элементов и связей между ними, архитектоника изучает архитектуру, рассматривает те же элементы как неповторяющиеся, как особенные по своему расположению и взаимной связи частей целого» (В. Н. Беклемишев, 1925. Морфологическая проблема животных структур). Гипотеза Грегори, даже будучи доказанной, способна решить проблему происхождения рыбьего плавника на тектологическом уровне, но не на архитектоническом. О месте плавников в плане строения рыб она не говорит почти ничего. Неясно, верна ли теория боковой складки (Грегори был обоснованно осторожен в этом пункте). Не будем забывать, что, несмотря на множество косвенных доводов, ни одного прямого подтверждения теория боковой складки до сих пор не имеет.
Плавники и гены
Теперь еще раз обратимся к биологии развития. В 1925 году киевский эмбриолог Борис Иванович Балинский продемонстрировал впечатляющий пример явления, которое называется эмбриональной индукцией. Пересадив под кожу на боку личинки тритона зачаток слухового пузырька, он обнаружил, что в этом месте развивается дополнительная — пятая — конечность (см., например: B. I. Balinsky, 1956. A new theory of limb induction). Дальнейшие опыты Балинского показали, что по боковой стороне тела у личинок или зародышей наземных позвоночных проходит узкая и длинная компетентная зона, готовая образовать конечность в ответ на действие индуктора (таковым может послужить что угодно — не только живая ткань, но даже кусочек целлоидина).
Японские биологи экспериментально показали, что у зародышей хрящевых рыб вдоль бока тоже проходит компетентная зона, действие на которую определенным FGF-белком вызывает там экспрессию генов, характерных для развивающихся конечностей (в том числе гена Shh). Казалось бы, чем не подтверждение теории боковой складки? Однако полностью непрерывной эта компетентная зона оказалась только у ската, гигантские грудные плавники которого перекрываются с брюшными. У акулы экспрессия генов конечностей появляется там не во всех тканях, а морфологически выраженные зачатки плавников — такие, как апикальный эктодермальный гребень — не образуются вообще. Такой результат выглядит, признаться, двусмысленно. Боковая компетентная зона акулы может быть остатком единой складки, но может и не быть. С тем же успехом это может быть чисто эмбриональный признак, связанный, например, с когда-то имевшим место перемещением плавников вдоль оси тела.
Кроме того, у водных позвоночных и у личинок амфибий есть еще одна компетентная зона — непарная, проходящая по средней линии спины и вокруг кончика хвоста к анальному отверстию. Оказалось, что в этой зоне можно индуцировать образование апикального эктодермального гребня, воздействуя FGF-белками. Более того, «пробудить» эту зону, вызвав в ней экспрессию соответствующих генов, можно даже у зародыша курицы.
В итоге японские исследователи констатируют, что данные генетики развития совместимы и с теорией боковой складки, и с теорией первичности грудного плавника, и с теорией первичности брюшного плавника, и даже с теорией происхождения парных плавников от непарных (рис. 11; S. Yonei-Tamura et al., 2008. Competent stripes for diverse positions of limbs/fins in gnathostome embryos). Для того, чтобы сделать между этими теориями выбор, нужны другие источники информации.
Многообразие древних плавников
Вновь обратимся к палеонтологии. Взглянем повнимательнее на акантод, тех самых вымерших рыб, у которых парные плавники всегда имеют шипы (а это, как мы знаем, очень древний признак) и часто присутствуют в числе нескольких пар. Эта группа изучена весьма неплохо — ей, например, еще в первой половине XX века посвятил серию работ выдающийся британский палеонтолог Дэвид Уотсон (David Meredith Seares Watson). И сейчас исследования акантод вовсю продолжаются. Нельзя не добавить, что многие современные палеонтологи считают акантод парафилитической группой, то есть скорее эволюционным уровнем, нежели ветвью. Не исключается, что среди акантод были предки как хрящевых, так и костных рыб (P. Ahlberg, 2009. Birth of the jawed vertebrates). Поэтому само название «акантоды» в современных статьях часто пишут в кавычках. Но эти систематические тонкости не влияют на интересующий нас предмет. Акантоды — точно очень древние и примитивные существа. С другой стороны, чего-чего, а парных плавников у них уже хватало. Не могут ли они дать какую-нибудь подсказку, касающуюся происхождения этих структур?
Чтобы ответить на этот вопрос, присмотримся к рисункам того же Уотсона, на которых в деталях изображены типичные акантоды (рис. 12; D. M. S. Watson, 1937. The Acanthodian fishes). Как правило, они имеют одну пару сильных грудных плавников и несколько пар более слабых брюшных. Все брюшные плавники одной стороны лежат на одной линии. Но вот грудные плавники на продолжении этой линии не находятся — они как бы расставлены пошире. Линия брюшных плавников проходит четко внутри от пары грудных. Разумеется, это несколько противоречит теории боковой складки, которая требует, чтобы грудные плавники лежали на одной линии с брюшными.
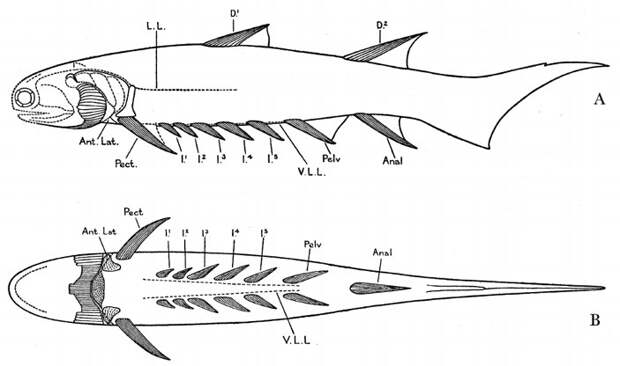
Брюшные плавники у акантод так сильно смещены внутрь, что могут даже сливаться посредине. Непарный брюшной плавник является диагностическим признаком одного из самых известных семейств акантод — семейства Acanthodidae (J. Zajic, 1995. Some consequences of recent investigations on the family Acanthodidae Huxley, 1861). Это относительно позднее семейство, оно появляется в карбоне. При этом у акантодид вперед от непарного брюшного плавника идут парные каналы боковой линии, что наглядно свидетельствует о вторичности непарного состояния (P. A. Beznosov, 2009. A redescription of the Early Carboniferous acanthodian Acanthodes lopatini Rohon, 1889).
У раннедевонской хрящевой рыбы Kathemacanthus (которую долго принимали за акантоду; G. Hanke, M. Wilson, 2010. The putative stem-group chondrichthyans Kathemacanthus and Seretolepis from the Lower Devonian MOTH locality, Mackenzie Mountains, Canada) грудных плавников тоже несколько пар. При этом ряды грудных и брюшных плавников у нее расположены под таким углом друг к другу, что сериальная гомология между ними исключена (рис. 13а; P. Gagnier, M. Wilson, 1996. Early Devonian acanthodians from northern Canada). Очевидно, она есть только внутри каждого из рядов. Получается, что грудные и брюшные плавники примитивных рыб — это негомологичные структуры.
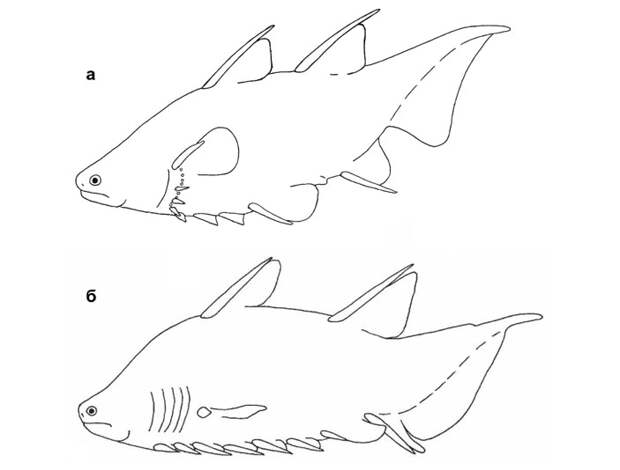
У раннедевонской акантоды Brochoadmones грудной плавник расположен прямо над серией из семи брюшных (рис. 13б; G. Hanke, M. Wilson, 2006. Anatomy of the Early Devonian acanthodian Brochoadmones milesi based on nearly complete body fossils, with comments on the evolution and development of paired fins). Само собой ясно, что в эту серию он не входит.
Канадские палеоихтиологи Гевин Ханке (Gavin Hanke) и Марк Уилсон (Mark Wilson) пишут, что многие раннедевонские челюстноротые имели «набор грудных, предгрудных, брюшных и предбрюшных плавников», причем все эти плавники были с шипами (G. Hanke, M. Wilson, 2005. Fin spines, scales, teeth, and prediction of early jawed fish structure). Таким образом, плавники древнейших рыб оказались гораздо многообразнее, чем можно было бы подумать.
Картина еще больше усложнится, если спуститься на один эволюционный этаж ниже и погрузиться в мир древних бесчелюстных. У остеострак, которых многие специалисты считают ближайшими родственниками рыб, единственные парные плавники расположены прямо над жаберным аппаратом. У телодонтов аналоги грудных плавников тоже находятся над рядом жаберных щелей. Впрочем, у силурийского телодонта Shielia есть еще и вторые парные плавники (аналоги брюшных), и они расположены явно ниже уровня жабр (рис. 14). А вот у анаспид парные плавники всегда находятся ниже ряда жаберных щелей. Их сериальная гомология с парными плавниками телодонтов весьма сомнительна.

Как же найти путеводную нить в этом хаосе?
Пять меридианов
В 2012 году Александр Николаевич Кузнецов опубликовал статью под названием «Пять долгот в теле хордовых» (A. N. Kuznetsov, 2012. Five longitudes in chordate body). В ней ставится вопрос о фундаментальных характеристиках тела позвоночных животных, которые можно было бы противопоставить устаревшему архетипу, предложенному в середине XIX века Ричардом Оуэном (Richard Owen). В этой работе Кузнецов не стал рассматривать голову позвоночных, которая имеет собственную сложную систему внутренней сегментации (см., например: С. А. Ястребов, 2018. Метамерия головы позвоночных: современное состояние проблемы). Он ограничился туловищем, попытавшись выделить моменты, которые исследователи часто упускали из виду.
Тело позвоночного (как и любого вторичноротого) имеет древние передний и задний концы, которые можно сравнить с северным и южным полюсами глобуса. «Северный полюс», то есть морфологический передний конец тела позвоночного, находится в области головы — мы сейчас можем не уточнять, где именно (это достаточно сложный вопрос, по которому высказывались разные мнения; см., например: G. de Beer, 1931. On the nature of the trabecula cranii). «Южный полюс», то есть морфологический задний конец тела, — это анальное отверстие, расположенное приблизительно на месте первичного эмбрионального рта — бластопора. У позвоночных спинное «полушарие» дало большой вырост назад за счет образования хвоста (который целиком расположен позади анального отверстия и не содержит полости тела), но топологически это ничего не изменило.
Гипотеза Кузнецова заключается в том, что в теле позвоночных есть пять устойчивых линий эктодермально-мезодермальных взаимодействий, то есть взаимодействий между внешним зародышевым листком — эктодермой — и лежащей под ним мезодермой (рис. 15). Это непарный срединный меридиан, парные боковые меридианы и парные млечные меридианы. Точкой, где все они сходятся, является анальное отверстие. Срединный меридиан проходит в основном по спинной стороне, но огибает хвост снизу. Боковые и млечные меридианы расходятся от анального отверстия по бокам туловища. Принципиальная разница между ними становится заметна, когда они достигают жаберного аппарата. Боковой меридиан проходит над жаберными щелями, а млечный меридиан — под ними. Кроме того, сразу позади последней жаберной щели меридианы пересекаются широтным клейтральным кольцом, под которым формируется плечевой пояс. Пожалуй, клейтральное кольцо можно было бы с тем же успехом назвать клавикулярным: клейтрум — это покровная кость плечевого пояса, расположенная выше плечевого сустава, а ключица (clavicula) — ниже сустава, куда клейтральное кольцо, разумеется, тоже заходит (см. рис. 5E в статье E. Jarvik, 1965. On the origin of girdles and paired fins).
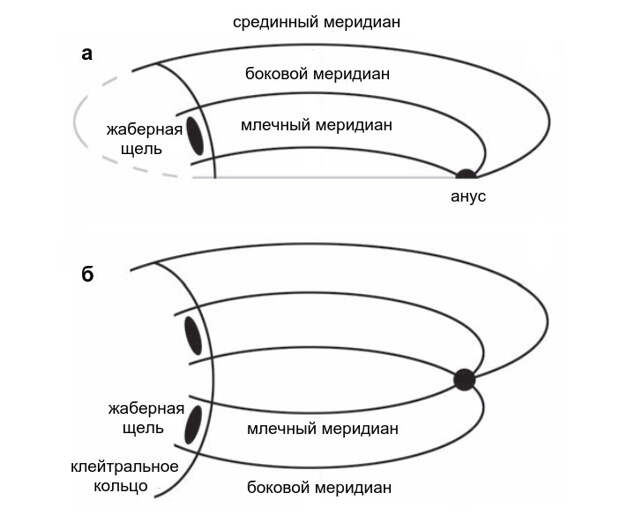
Эктодермально-мезодермальные взаимодействия в развивающемся организме позвоночных многообразны. Эктодерма в местах специфического контакта с мезодермой может впячиваться, выпячиваться, формировать железы, чешуи, шипы, подкожные каналы (в случае с боковой линией) или же «запускать» посредством апикального эктодермального гребня развитие зачатка конечности, в который заходят мышечные почки и нервные окончания всех ближайших к нему сегментов тела. Из эктодермы образуются покровы и нервные элементы, из мезодермы — мышцы и скелет конечности. Выделенные Кузнецовым меридианы — это узкие вытянутые вдоль тела компетентные зоны, где экспрессия генов, обеспечивающих каскад взаимодействий между эктодермой и мезодермой, сильно облегчена.
Как спинной, так и анальный плавники находятся на срединном меридиане. На его непосредственном продолжении, то есть на средней линии тела впереди от анального отверстия, непарные эктодермально-мезодермальные структуры обычно не формируются, хотя чисто физиологически этому вроде бы ничто не мешает. Парные структуры, возникшие на млечном меридиане, изредка сливаются посредине (как брюшные плавники акантодид), но в таких случаях всегда есть признаки того, что непарное состояние вторично.
На основе архетипа Кузнецова можно сделать несколько выводов. Прежде всего, меридианы эктодермально-мезодермальных взаимодействий — это чисто эмбриональная адаптация. Никаких непрерывных боковых складок у взрослых животных, скорее всего, ни на каком эволюционном этапе не было. Но были устойчивые компетентные линии, в любой точке которых можно было заложить плавник.
Кроме того, новый архетип позволяет худо-бедно навести порядок в хаосе всевозможных парных плавников (рис. 16). По боковому меридиану возникли единственные парные плавники остеострак, передние парные плавники телодонтов и, вероятно, метаплевральные складки ланцетника, которые появляются у его личинки, а у взрослого ланцетника срастаются под брюхом и замыкают атриальную (околожаберную) полость. Некоторые авторы считали метаплевральные складки ланцетника предшественниками тех самых боковых складок, из которых у позвоночных образовались парные плавники. Но только что рассмотренные палеонтологические данные убеждают нас, что на самом деле все сложнее.
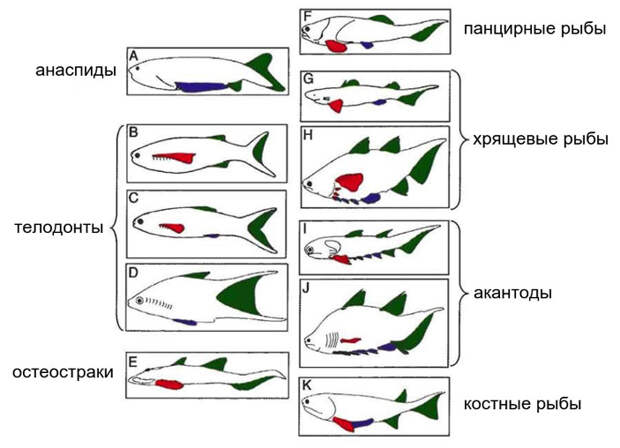
По млечному меридиану возникли задние парные плавники телодонтов, все парные плавники анаспид, брюшные плавники рыб, а также развившиеся отдельно от брюшных плавников совокупительные органы вымерших панцирных рыб, которые, похоже, были общим признаком этой древней группы (см. J. A. Long et al., 2015. Copulation in antiarch placoderms and the origin of gnathostome internal fertilization).
Грудные плавники рыб всегда расположены на клейтральном кольце, сразу за жабрами. Может быть, они здесь и возникли? Именно зону клейтрального кольца занимает ряд из нескольких грудных плавников у древней хрящевой рыбы Kathemacanthus (см. рис. 13а). Если же грудные плавники сильно разрастаются, как, например, у скатов, то они могут распространиться на боковой меридиан. Вот почему грудной плавник ската перекрывает брюшной именно сверху.
Не исключено, однако, что грудные плавники рыб возникли на млечном меридиане и только потом постепенно «переползли» на клейтральное кольцо, заняв в итоге свое нынешнее место позади жабр. Если это так, то теория первичности брюшных плавников верна. Но считать это доказанным пока нельзя.
«Пять долгот тела хордовых» служат основой не только для плавников. Структура, наиболее характерная для бокового меридиана рыб — это боковая линия, которая, как правило, тянется на большую часть туловища (у миног и миксин сплошной боковой линии нет). По млечному меридиану у млекопитающих обычно закладываются ряды молочных желез.
Вполне вероятно, что примитивные позвоночные, никогда не имевшие парных конечностей (хайкоуихтис, гетеростраки, предки миног и миксин), обладали только срединным меридианом, а другие меридианы возникли позже путем его неоднократной дупликации. Как раз такую последовательность событий, но более упрощенную, предполагала уже знакомая нам Зерина Джохансон. Таким образом, теория происхождения парных плавников из непарных выглядит очень правдоподобно, хотя это и не противоречит ни теории плавниковых шипов Грегори, ни (возможно!) теории первичности брюшных плавников. Но даже если последняя верна, это касается только рыб: у бесчелюстных пути эволюции могли быть иными.
Свойственное костным рыбам расположение брюшных плавников внутри от грудных — очень древний признак. Именно сохранение этого признака позволяет брюшным плавникам в некоторых случаях «выезжать» вперед от грудных — например, у современных трескообразных рыб (см. изображение налима). Наоборот, расположение грудных и брюшных конечностей примерно на одной линии, свойственное, с одной стороны, акулам, а с другой стороны — наземным позвоночным, есть независимо возникшая в этих группах поздняя черта.
Преанальный плавник, или еще раз о происхождении парности
А вот теперь обсудим вышедшую летом 2023 года в журнале Nature статью по биологии развития, подписанную биологами из Сингапура, США, Швейцарии, Чехии, Англии и Австралии. Ее главным героем стала аквариумная рыбка данио-рерио (Danio rerio), излюбленный объект эмбриологов, изучающих водных позвоночных. В научном сообществе эту полосатую рыбку называют «зебрафиш» (zebrafish).
Заинтересовавшись эмбриологическими данными, способными так или иначе прояснить происхождение парных плавников, авторы обратили внимание вот на какой интересный факт. Личинка данио-рерио, как и положено, имеет непарную плавниковую складку, которая проходит по спине и огибает хвост. Из этой складки образуются спинной, хвостовой и анальный плавники. Но, кроме того, у личинки данио есть дополнительная непарная складка, расположенная на средней линии брюха впереди от анального отверстия. Она называется малой лопастью или преанальной плавниковой складкой (pre-anal fin fold, PAFF).
В ходе метаморфоза преанальная плавниковая складка исчезает. Никакие структуры взрослого животного из нее не образуются. Но откуда она вообще взялась? И куда потом делась, а точнее — какую роль сыграла в эволюции?
Прежде всего исследователи решили выяснить, откуда в преанальной складке берется зародышевая соединительная ткань — мезенхима. В плавниках она, как правило, имеет мезодермальное происхождение. Но сама мезодерма у позвоночных устроена очень сложно. Сейчас достаточно сказать, что она делится на два региона: параксиальная мезодерма (преимущественно спинная) и мезодерма боковой пластинки (преимущественно брюшная). Из боковой пластинки образуется много разных структур, в том числе миокард, мышцы глотки, целомическая выстилка и — что нам сейчас особенно важно — скелет парных плавников (см. Ф. Я. Дзержинский, 2005. Сравнительная анатомия позвоночных животных).
Параксиальная мезодерма и боковая пластинка отличаются друг от друга на уровне экспрессии некоторых генов. Например, существует особый белок, который синтезируется в параксиальной мезодерме, но не в мезодерме боковой пластинки. Проверив наличие этого белка у личинки данио, авторы увидели, что в главной непарной плавниковой складке он есть, а вот в преанальной — нет. Зато в преанальной плавниковой складке экспрессируется регуляторный ген Hand2, активность которого характерна именно для структур, развивающихся из боковой пластинки (рис. 17). В главной непарной плавниковой складке экспрессия этого гена, как и ожидалось, отсутствует. Наконец, исследователи пометили клетки боковой пластинки у эмбриона, заставив их синтезировать определенный трансгенный белок, и обнаружили этот белок в мезодерме преанальной плавниковой складки личинки. Попасть туда он мог только вместе с мигрирующими клетками. Итак, мезодермальная «начинка» преанальной плавниковой складки, несомненно, происходит из боковой пластинки — как и скелет парных плавников.
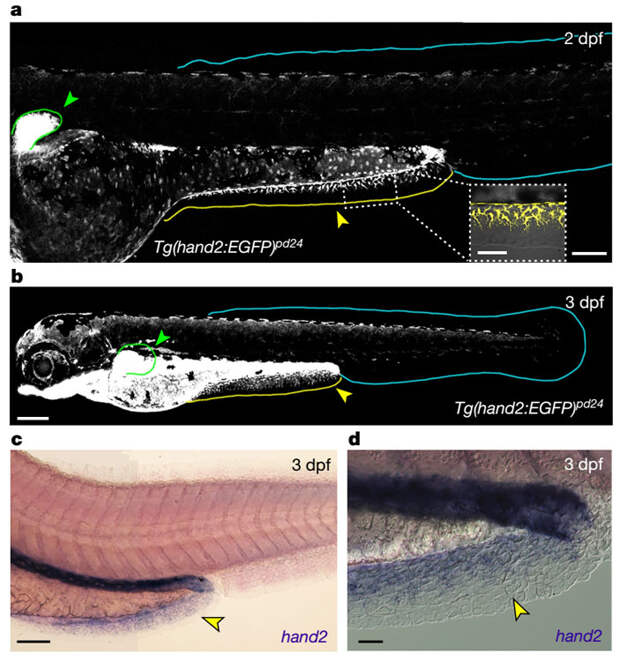
Что это — уникальная особенность данио или древний признак, общий с другими рыбами? Скорее всего — второе. Это показывает сравнительное исследование. У личинки японской медаки преанальная плавниковая складка невелика, но все-таки выражена, и экспрессия гена Hand2 в ней, в отличие от главной плавниковой складки, есть. У американского веслоноса, представителя одной из самых древних и архаичных групп костных рыб — осетрообразных (см. Китайский веслонос окончательно признан вымершим, «Элементы», 02.08.2022), на личиночной стадии тоже есть преанальная плавниковая складка, в которой тоже наблюдается экспрессия Hand2. И даже у личинки миноги, которая вообще рыбой не является, есть маленькая преанальная плавниковая складочка, в которой экспрессируется миножий гомолог гена Hand2 (он называется HandA). Правда, у миноги этот ген почему-то экспрессируется и в зачатке спинного плавника. Наконец, небольшая преанальная плавниковая складка обнаружена у зародышей австралийской двоякодышащей рыбы рогозуба и у головастиков шпорцевых лягушек, причем в последнем случае удалось показать, что в этой складке опять-таки экспрессируется ген Hand2. Напомним, что экспрессия этого гена, как и развитие скелета из боковой пластинки мезодермы — общая черта преанальной плавниковой складки с парными плавниками. И похоже, что наличие этой складки — очень древний признак позвоночных.
Само по себе это не новое открытие. Сравнительные анатомы еще сто лет назад знали о существовании преанальной плавниковой складки. Эдвин Гудрич (Edwin Stephen Goodrich) писал, что преанальная складка наблюдается в развитии панцирной щуки, амии и многих костистых рыб (E. S. Goodrich, 1930. Studies on the structure & development of vertebrates). Более того, некое подобие преанального плавника есть у миксины (рис. 18). Наконец, у некоторых родов ланцетников правая метаплевральная складка может продолжаться по средней линии тела вперед от анального отверстия, тем самым становясь непарной.
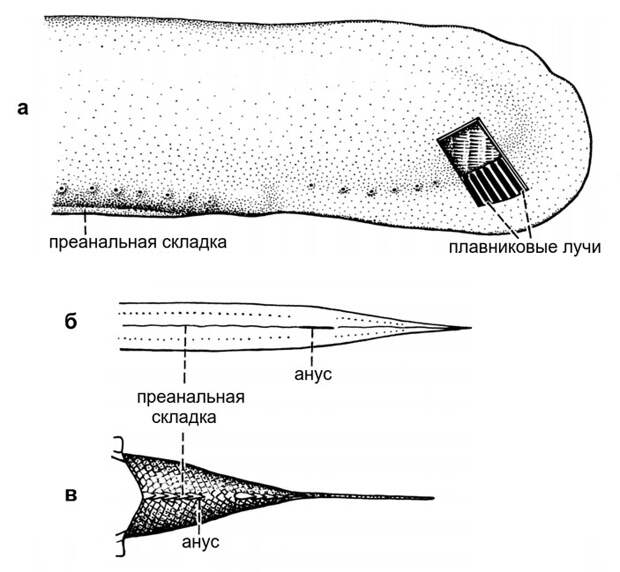
Правда, преанальную плавниковую складку не удалось обнаружить у зародышей самых примитивных современных рыб — акул. Или в линии хрящевых рыб она исчезла, или же костные рыбы и бесчелюстные «изобрели» ее независимо друг от друга. Авторы обсуждаемой статьи считают более правдоподобным первый вариант.
А нельзя ли удвоить преанальную плавниковую складку, чтобы она стала парной? Оказывается, можно. Известно, что подобные уродства иногда возникают у рыб в результате нарушения работы регуляторного гена, который называется Chordin. Ген этот знаменит тем, что у всех двусторонне-симметричных животных, кроме хордовых, он экспрессируется на брюшной стороне зародыша, и только у хордовых — на спинной (см., например: C. Lowe et al., 2006. Dorsoventral patterning in hemichordates: insights into early chordate evolution). Белок Chordin служит антагонистом белков группы BMP (BMP4, BMP7), которые, наоборот, маркируют брюшную сторону у зародыша хордовых и спинную — у всех остальных двусторонне-симметричных животных. В нашем случае исследователи ввели личинке данио небольшую дозу специальных синтетических молекул, блокирующих работу гена Chordin (см. Морфолино), и получили в ответ «вентрализацию», то есть разрастание брюшной стороны и усиление ее признаков. Вот при этой «вентрализации» как раз и возникло удвоение преанальной плавниковой складки, которая стала в результате парной и симметричной. Здесь невозможно не вспомнить прозвучавшее больше 150 лет назад высказывание доктора Джорджа Хамфри: парные плавники рыб — гомологи непарного анального плавника, которым помешал слиться внутренностный мешок. Если рассматривать зародышевый преанальный плавник как продолжение анального (а почему бы нет?), все именно так и выглядит.
Удвоение преанального плавника в целом не нарушает жизнеспособности рыбы, а значит, такая вариация вполне могла когда-то в далеком прошлом стать полноценным объектом естественного отбора. Правда, ни у каких известных позвоночных — ни у современных, ни у ископаемых — нет во взрослом состоянии непарного преанального плавника (видимо, за исключением миксин и их родственников). Позвоночные быстро «проскочили» этот эволюционный этап либо никогда его толком и не проходили: не исключено, что преанальный плавник всегда был преимущественно личиночным органом. Однако его удвоение (которое изначально могло быть случайным) сразу создало структуру, оказавшуюся полезной во взрослом состоянии: пару стабилизирующих боковых килей. Свои функции они выполняли несовершенно, но в условиях, когда парных плавников не было ни у кого, преимуществом было и это.
Так, вероятно, и возник млечный меридиан Кузнецова. Утверждение, что на средней линии брюшной стороны у позвоночных никогда не формируются эктодермально-мезодермальные структуры, надо признать поспешным: его опровергает преанальная плавниковая складка. Но первично непарных органов взрослых животных на этой линии, как правило, нет. Зато она породила млечный меридиан, на котором — если авторы обсуждаемой статьи правы — у позвоночных и образовались первые парные плавники.
Иллюстрацией этого предполагаемого эволюционного этапа могут послужить ископаемые бесчелюстные из группы анаспид. Их вытянутые лентовидные брюшные плавники очень похожи на то, что должно было бы образоваться в результате расщепления непарного преанального плавника. В свою очередь, брюшные плавники анаспид прошли собственную эволюцию: они могли вовсе не иметь твердого скелета (у Jamoytius; см. R. Sansom et al., 2010. Taphonomy and affinity of an enigmatic Silurian vertebrate, Jamoytius kerwoodi White) или иметь внутренний скелет, состоявший из параллельных радиалий.
Совсем недавно «вентролатеральные плавниковые складки», похожие на брюшные плавники анаспид, были описаны у представителя другой группы древних бесчелюстных, а именно галеаспид (см. Панцирное бесчелюстное туцзяаспис проливает свет на происхождение конечностей позвоночных, «Элементы», 10.10.2022). Кроме того, подобные «вентролатеральные складки» есть и у остеострак, которые морфологически довольно близки к галеаспидам.
И вот здесь мы упираемся в проблему происхождения грудных плавников. Ведь остеостраки тем и выделяются в мире древних бесчелюстных, что у них есть отлично развитые грудные плавники. Правда, они устроены совершенно не так, как грудные плавники рыб, и гомология здесь сомнительна (см. Л. И. Новицкая, В. Н. Каратаюте-Талимаа, 1986. Замечания о кладистическом анализе в связи с гипотезой миоптеригий и проблемой происхождения гнатостом). Но в любом случае — откуда они взялись? Плоские горизонтальные грудные плавники остеострак занимают однозначно наджаберное положение, в то время как и у анаспид, и у галеаспид, и у самих остеострак брюшные плавниковые складки пролегают ниже уровня жаберных щелей. К тому же у остеострак эти складки до жабр не доходят. Таким образом, между грудными и брюшными плавниками изначально — с момента их появления в палеонтологической летописи — имеется разрыв. В терминах Кузнецова брюшные плавниковые складки находятся на млечном меридиане, а грудные плавники остеострак — на боковом. Простым продолжением вперед линии брюшных плавников грудные плавники не получить!
С этой точки зрения крайне интересен представитель анаспид под названием Rhyncholepis, на которого авторы обсуждаемой работы справедливо обратили особое внимание (рис. 19; A. Ritchie, 1980. The Late Silurian anaspid genus Rhyncholepis from Oesel, Estonia, and Ringerike, Norway). У ринхолеписа есть типичные для анаспид брюшные плавниковые складки. Но впереди они переходят в крупные треугольные плавники, устроенные довольно примечательным образом. Во-первых, передний край плавника снабжен самым настоящим твердым и острым шипом — вспомним теорию Грегори! Во-вторых, основание этого шипа охватывает последнюю жаберную щель сзади, заходя длинными отростками как ниже, так и выше нее. По похожему плану устроен и еще один представитель анаспид — Pharyngolepis. Это тот самый момент «переползания» потенциального грудного плавника с млечного меридиана на клейтральное кольцо, о котором можно было догадаться, исходя из гипотезы Кузнецова.

Источник: Keh-Weei Tzung, Robert L. Lalonde, Karin D. Prummel, Harsha Mahabaleshwar, Hannah R. Moran, Jan Stundl, Amanda N. Cass, Yao Le, Robert Lea, Karel Dorey, Monika J. Tomecka, Changqing Zhang, Eline C. Brombacher, William T. White, Henry H. Roehl, Frank J. Tulenko, Christoph Winkler, Peter D. Currie, Enrique Amaya, Marcus C. Davis, Marianne E. Bronner, Christian Mosimann and Tom J. Carney. A median fin derived from the lateral plate mesoderm and the origin of paired fins // Nature. 2023. DOI: 10.1038/s41586-023-06100-w.
Сергей Ястребов
Свежие комментарии