
Нейробиологи из Англии, США и Канады применили методы работы с искусственными нейросетями к анализу большого массива реальных данных по активации нейронов мозга. Это методики по выявлению скрытой динамики нейроактиваций в больших массивах разнородных данных с техническим шумом. Ученые показали, что то или иное сложное движение управляется популяцией нейронов со встроенной единообразной картиной активаций. Эта картина выявляется во всех тестах у одного животного, а также в схожих тестах у разных животных. Более того, единообразные активации найдены не только при выполнении самого действия, но и при его планировании. На моделях ученые доказали, что выявленную единообразную картину нельзя объяснить только сходством самих действий. Требуются дополнительные гипотезы. Авторы предполагают, что популяции нейронов с единообразной картиной активаций являются функциональными компартментами, управляющими обучением тому или иному действию и его исполнением.
Всем известно, что в мозге животных запредельно много нейронов: по грубым прикидкам в человеческом мозге их около 85 миллиардов и даже у рыбки данио-рерио — порядка десяти миллионов. Несмотря на такое разнообразие, при решении одной и той же задачи у людей естественно ожидать определенного поведения: каждый у входной двери нажмет на кнопку звонка, школьники встанут со стула, когда входит директор, все посмотрят по сторонам при переходе улицы, и в других типовых ситуациях большинство выполняет одно и то же условно-рефлекторное действие. С животными аналогично: обученные рыбки поплывут к кормушке при определенном звуковом сигнале, обученные мыши в нужной ситуации нажмут на рычаг и т.
д. Иными словами, при конкретном запросе все так или иначе приходят к одному и тому же решению.С учетом миллионов нейронов, вовлеченных в контроль каждого действия (даже у рыбки данио при рефлекторном движении к кормушке активируется около 500 000 нейронов), уместно задать вопрос, как устроен контроль каждого сложного действия. В настоящее время при осмыслении этого вопроса привлекаются две модели: шеррингтоновская и модель Хопфилда (рис. 1).
Шеррингтоновская модель, которая разрабатывалась в основном с середины прошлого века, дает представление о работе мозга на основе сложных каскадов передачи нервных импульсов (см. статью Обратная сторона рефлекса). Эти каскады формируют сложные сети и петли обратных связей, позволяющие координировать действия разных мышц. При обучении должны формироваться новые синапсы и шипики на нейронах, и таким образом новые пути активации перестраивают имеющиеся каскады. Шеррингтоновская модель плохо справляется с большим числом измерений, когда снимаются данные не с десятка нейронов, а с сотен и тысяч. А именно работа с гигантскими ансамблями данных сейчас в ходу у нейробиологов. Когда данных много, то картина передачи сигналов даже в одном небольшом участке мозга размывается, общий тренд тонет, неуловимый среди хаоса нейронных активаций и технического шума. Каскады передачи сигналов больше не выявляются.
Ученые предположили, что если хочется понять нейробиологию поведения, то следует отказаться от поиска цепочки передачи сигналов от нейрона к нейрону: слишком сложно осмыслить и нарисовать рабочую схему из тысячи узлов и связей. Вместо этого стоит попробовать понять целостную картину активаций всех нейронов того или иного участка мозга. То есть работать с хопфилдовскими нейросетями. В них каждая связь между каждой парой нейронов изначально признается равноценной, при обучении меняется восприимчивость к поступающим сигналам в тех или иных нейронах, таким образом создается поток сигналов для выполнения конкретного мышечного движения. Здесь в меньшей степени важно, как работают синапсы и куда в них передается сигнал, а в большей степени важно, как данная сеть настраивается (обучается). Такой подход имеет глубокий смысл, так как, описывая контроль того или иного механического действия, нейробиологи оперируют не столько отдельными нейронами, сколько целыми нейронными ансамблями или даже участками мозга.
Однако, как работать с отдельными нейронами и цепочками передачи сигналов, приблизительно ясно: нужно снимать микроэлектродами потоки импульсов в нейронах. А как работать с популяциями и сетями реальных нейронов? Ведь эти нейроны не запрограммированы в искусственной нейросети, а реальные, в живом мозге. Такая задача похожа на виток спирали: сначала поняли, что существуют нейроны и по ним передается сигнал, потом научились математически моделировать передачу сигналов в сложных сетях (нейросети), а теперь нужно наработанные принципы обращения с нейросетями приложить к реальному мозгу.
В недавнем выпуске журнала Nature опубликована статья, авторы которой описывают, во-первых, как это делать, и, во-вторых, какие результаты дает такой подход. Данная работа выполнена в Лаборатории биоинженерии Лондонского королевского колледжа под руководством Хуана Галлего (Juan Alvaro Gallego) и Мэттью Перича (Matthew G. Perich). Ученые, в том числе и руководители данной работы, уже около десяти лет занимаются разработкой методик анализа реальных нейронных показателей с позиций математических нейросетей.
В их лаборатории работают с обезьянами и мышами. В мозг подопытных животных (в моторную и премоторную кору и базальные ганглии) вживлены микрочипы, с которых можно снять потенциалы нескольких тысяч нейронов из двух-трех отделов мозга. Животных можно обучать тому или иному действию, а потом отследить картину нейронных активаций во время выполнения данного действия до и после обучения, а также проследить весь процесс обучения. Обезьян обучали двигать курсор на экране компьютера от точки в центре к точке на периферии. Точки могли быть разных цветов и находиться в разных частях экрана. Мышей учили нажимать на рычаг определенного веса (тяжелый и легкий): нажмешь не с той силой — не получишь награду. В результате поведенческих экспериментов с микрочипов собирали данные по динамике активаций каждого нейрона трех участков мозга. С таким количеством данных методики по анализу нейронных каскадов не применимы, здесь подойдет только модель нейросети. Именно методами анализа нейросетей выявляется общий знаменатель всех активаций. Для математиков такая задача будет формулироваться как снижение размерности пространства: требуется многомерное пространство связей каждого нейрона с каждым превратить в пространство с малым числом измерений (проекций). Это малое число измерений и будет направлением движения сигнала, поступившего в тот или иной участок мозга, к двигательным мышцам. Простейший подход к решению такой задачи — метод главных компонент. Здесь, конечно, полезно поразбираться с методикой (см. дополнительные материалы к статье), так как нейробиологам, работающим с колоссальными массивами данных, необходимо уметь выявлять из хаоса измерений основную тему.
Как бы ни был сложен математический аппарат подобного анализа, он позволяет в большом реальном массиве возбуждений нейронов выделить единую траекторию активаций во время выполнения конкретного действия. Эта единая траектория называется в англоязычной литературе latent dynamics. В русскоязычной научной литературе по нейробиологии аналога этого термина нет (или, скорее всего, мне не известна такая литература); здесь я буду использовать словосочетание «скрытая траектория», подразумевая, что общая траектория активаций скрыта в массиве множественных разнородных и разнокачественных данных.
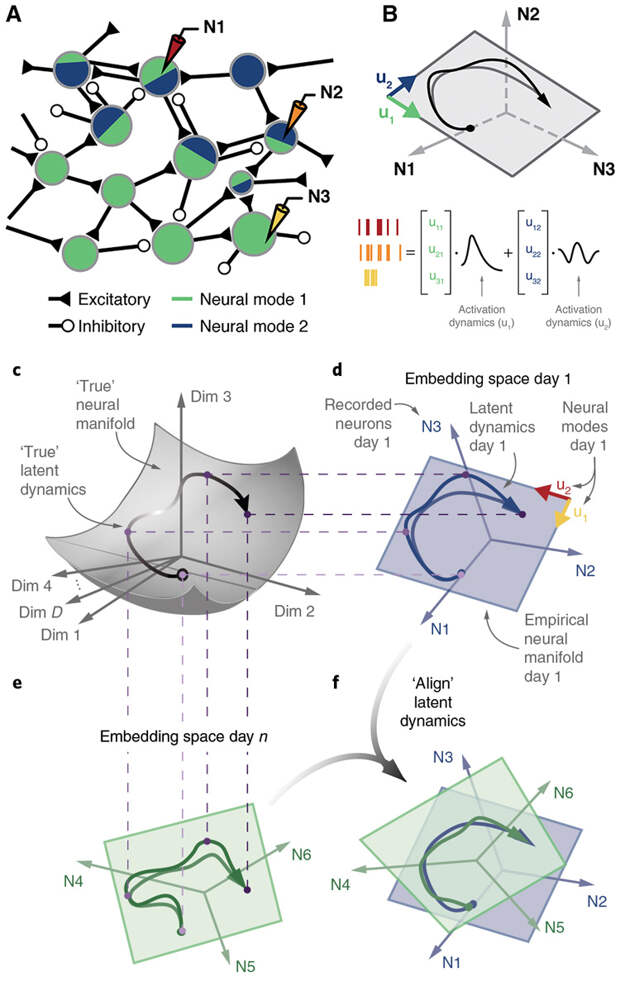
В обсуждаемой статьи анализировались массивы данных по активации нейронов у трех обезьян и четырех мышей. Каждое животное проходило тесты по двигательным упражнениям (двигать курсор на экране у обезьян и нажимать на рычажок к кормушке у мышей) по десять раз. После каждого теста выявлялась скрытая траектория активаций. Затем, после выравнивания, полученные скрытые траектории сравнивались для каждого животного: насколько они схожи? Затем сравнивали скрытые траектории между всеми обезьянками и между всеми мышами: найдется ли общее для всех них?
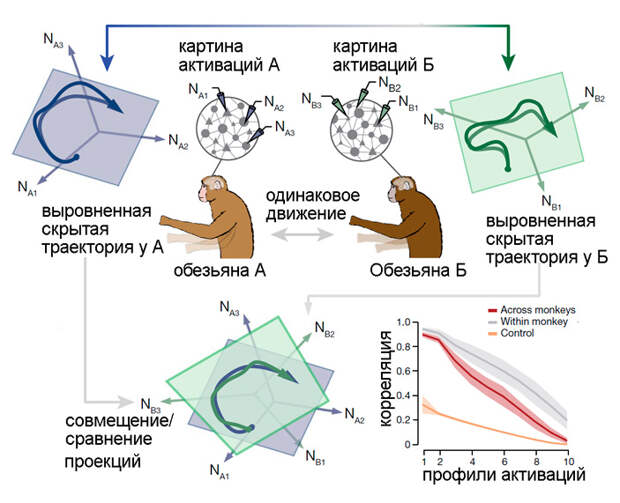
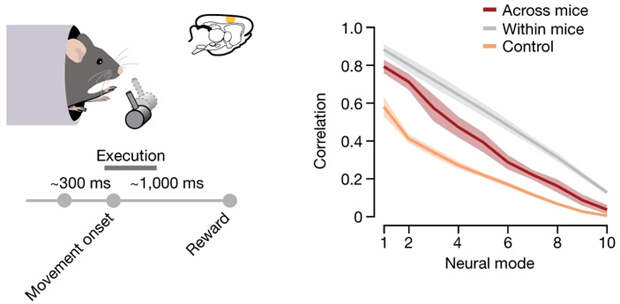
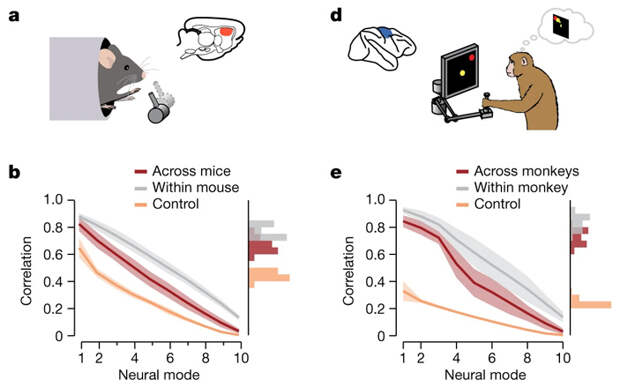
Оказалось, что для всех тестов у одного животного скрытая траектория моторной коры мозга (а точнее, участка с вживленными электродами) была сходной (рис. 3 и 4). Не меньшее сходство в скрытых траекториях выявилось и при сравнении тестов между животными (всех обезьян или всех мышей). Эта траектория для обезьян оказалась устойчивой к проверкам, так как в контрольной выборке, организованной из смеси имеющихся данных с сохранением статистической структуры активаций, траектория потерялась. В опытах с мышами выявились ровно такие же закономерности, правда уровень сходства как между тестами одного животного, так и между животными был ниже, чем у обезьян.
Научившись выявлять скрытую траекторию, ученые задались следующим вопросом: будет ли она выявляться в разных отделах мозга? Вполне вероятно, что процесс обучения движениям связан с моторной корой, особенностями ее архитектуры и проекций нейронов в этот отдел. По налаженной схеме был проведен анализ активности нейронов в дорзальной части полосатого тела — отдела конечного мозга, также связанного с организацией движений. И что же? Нейроны полосатого тела также активировались по общей скрытой траектории (рис. 5).
Другой важный вопрос, поставленный в обсуждаемой работе, — будет ли скрытая траектория выявляться при планировании того или иного движения? Для его решения эксперименты чуточку видоизменили. Обезьяны должны были выполнить движение курсором по экрану не сразу после появления точки на экране, а по звонку, который раздавался спустя некоторое небольшое время. В этот короткий временной промежуток животное готовилось выполнить движение. То есть мысленно планировало движение. В течение этого промежутка тоже велась запись активаций моторной коры. К радости экспериментаторов, скрытая траектория выявилась и в этом случае. Она напоминала ту, которая получалась при выполнении самого движения.
Как объяснить существование скрытых траекторий при выполнении выученных движений? Первое, что естественно приходит на ум, это сходство самих движений. Обезьян, как и мышей, специально учили делать нечто сходное, не удивительно, что для этих действий нужны сходные ансамбли нейронов. Проверить это тривиальное предположение не просто, потому что невозможно разобраться в хаосе взаимосвязей «по-нейронно». Но ученые все же постарались проверить эту гипотезу, выбрав путь моделирования нейросетей (это как раз у них хорошо налажено). Они выявили необходимый набор реальных сигналов на входе в мозг и на выходе, то есть поступающий уже к мышцам. Затем натренировали модельную нейронную сеть продуцировать те же сигналы при тех же параметрах входа (иными словами, с тем же входящим сигналом, приводящим к требуемому финальному движению). А потом посмотрели на модельные скрытые траектории в модельных нейросетях. И получили набор траекторий, отличных от реальных. Следовательно, сходство конечных движений необходимо, но не достаточно для получения сходной скрытой траектории активаций. А вот что нужно еще, ученые ответить не смогли. Пока не смогли. Они лишь предположили, что это сходство обеспечивается некоей внутренней организацией нейронных популяций в каждой зоне мозга. Эта организация определяется, как им кажется, генетически и внешним контекстом развития каждого индивидуума. В пределах популяций нейронов, ограниченных в своих действиях всеми этими факторами, происходит обучение тем или иным действиям.
Оправдаются эти предположения или нет, но результат этих исследований замечательный. Он показывает существование функциональных ансамблей нейронов, настроенных на обучение. Своего рода единицы когнитивной активности. Если вдруг окажется, что такие ансамбли со своей скрытой траекторией активации обслуживают не только отдельные моторные действия, но и другие функции, то это будет настоящий прорыв в нейробиологии, создание новой картины работы мозга. Ученые приблизятся к пониманию, как организуется сложное поведение. Причем не на уровне отдельных нейронов и их «эстафетных команд», и не крупных участков мозга, пусть даже со своей морфологией и архитектурой, а на каком-то среднем уровне, более пригодном для осмысления целостной динамики передачи и преобразовании информации в мозге.
Именно из-за этой перспективы я решила взяться за написание этой новости, трудной для понимания во всех отношениях. Надеюсь, однако, что даже в моем изложении данная работа окажется полезной российским специалистам.
Источник: Mostafa Safaie, Joanna C. Chang, Junchol Park, Lee E. Miller, Joshua T. Dudman, Matthew G. Perich & Juan A. Gallego. Preserved neural dynamics across animals performing similar behaviour // Nature. 2023. DOI: 10.1038/s41586-023-06714-0.
Елена Наймарк
Свежие комментарии