

Рис. 1. Некоторые представители животного царства, у которых встречается однополое сексуальное поведение. a — морской ёж Lytechinus variegatus, b — летучая лисица Pteropus pselaphon, c — брюхоногий моллюск Crepidula fornicata (морская туфелька), d — кальмар Гумбольдта (Dosidicus gigas), e — обыкновенная подвязочная змея (Thamnophis sirtalis), f — белый гусь (Anser caerulescens), g — стрекоза Ischnura elegans, h — темноспинный альбатрос (Phoebastria immutabilis), i — малый булавоусый хрущак (Tribolium castaneum), j — полевой сверчок Teleogryllus oceanicus, k — домашняя корова (Bos taurus), l — морская звезда Archaster angulatus, m — японский макак (Macaca fuscata), n — антарктический пингвин (Pygoscelis antarcticus), o — серая жаба (Bufo bufo), p — нематода Nippostrongylus brasiliensis, q — губан-чистильщик Labroides dimidiatus, r — краб Calappa sp. Изображение из обсуждаемой статьи в Nature Ecology & Evolution
Широкая распространенность в животном мире однополого сексуального поведения традиционно рассматривается как эволюционный парадокс. В основе большинства гипотез, предложенных для его объяснения, лежат два постулата: 1) о резко негативном влиянии однополого секса на приспособленность (отсюда — поиск важных прямых или косвенных преимуществ, перевешивающих ущерб) и 2) о том, что строго разнополый секс — эволюционно исходное состояние для животных, а однополый — позднейшая «надстройка». Провокативное эссе, опубликованное группой американских биологов в журнале Nature Ecology & Evolution, ставит оба постулата под сомнение. По мнению авторов, эволюционно исходным состоянием могло быть неразборчивое половое поведение, направленное на любых представителей своего вида и не требующее умения определять пол партнера, а негативное влияние эпизодического (необлигатного) однополого секса на репродуктивный успех часто невелико или даже вовсе отсутствует. В таком случае в специальных объяснениях нуждается не эпизодическое однополое, а строго разнополое сексуальное поведение, требующее развития сложных систем идентификации партнеров и сопряженное с риском упущенных репродуктивных возможностей.
Группа биологов из Калифорнийского университета в Беркли и ряда других американских университетов опубликовала в уважаемом научном журнале Nature Ecology & Evolution увлекательное эссе, предлагающее в корне пересмотреть устоявшиеся подходы к изучению эволюционных основ гомосексуального поведения. Для начала уточняется терминология: во избежание излишнего антропоморфизма авторы отказываются от термина «гомосексуальность» в пользу «однополого полового поведения» (same-sex sexual behaviours, SSB). SSB понимается как любое поведение, направленное на представителя своего пола, которое, будучи направленным на представителя противоположного пола, может способствовать успешному размножению. Соответственно, вместо «гетеросексуальности» авторы говорят о «разнополом половом поведении» (different sex sexual behaviours, DSB).
Если рассматривать половое поведение как средство максимизации репродуктивного успеха особи (а это вполне классический эволюционный подход), то широкая распространенность в животном мире вариантов полового поведения, отличных от строгого DSB, на первый взгляд выглядит как парадокс, нуждающийся в специальном объяснении. Между тем у животных половое поведение действительно сплошь и рядом бывает направлено не на готовых к размножению представителей противоположного пола, а на другие объекты: на самого себя (самостимуляция), на особей того же пола (SSB), на представителей других видов, на мёртвых особей и даже на неодушевленные предметы. Что касается собственно SSB, то оно задокументировано более чем у полутора тысяч видов животных, относящихся ко всем большим ветвям позвоночных и ко многим типам беспозвоночных.
Восемнадцать из этих полутора тысяч видов показаны на рис. 1. По каждому виду авторы приводят пояснения, как именно проявляется SSB у данного вида животных, со ссылками на соответствующие исследования. Например, самцы летучих лисиц (рис. 1, b) вылизывают друг другу эрегированные пенисы, причем это поведение не связано с аллогрумингом, который для данного вида не характерен. Брюхоногие моллюски морские туфельки (рис. 1, c) — «последовательные гермафродиты»: они начинают свою взрослую жизнь как самцы, а потом, подрастая, превращаются в самок. Мелкие, подвижные самцы стараются примкнуть к компании особей покрупнее (обычно это самки), но некоторые предпочитают держаться ближе к другим самцам. Самцы некоторых кальмаров (рис. 1, d) прикрепляют сперматофоры к самцам и самкам своего вида с одинаковой частотой (J. T. Hendrik et al., 2011. A shot in the dark: same-sex sexual behaviour in a deep-sea squid). Спаривание обыкновенных подвязочных змей (рис. 1, e) часто происходит в больших клубках, где множество самцов пытаются спариться с одной особью, в роли которой иногда выступает самка, а иногда — самец, выделяющий похожие феромоны. У белых гусей (рис. 1, f) часто наблюдаются как попытки самцов спариться друг с другом, так и устойчивые женские пары, совместно воспитывающие своих гусят. И так далее.
Нельзя не заметить, что авторы трактуют SSB довольно широко. Как человек, много лет изучавший морских ежей, я поначалу был крайне удивлен присутствием морского ежа (рис. 1, а) среди видов, замеченных в гомосексуальном поведении. Ведь морские ежи не спариваются и не ухаживают друг за другом. Возле макушки панциря у них находятся пять половых отверстий (генитальных пор), из которых самцы выметывают прямо вверх, в воду, свои сперматозоиды, а самки — яйцеклетки. Оплодотворение у них наружное, сперматозоиды сами, как могут, ищут в воде яйцеклетку. Половой диморфизм у морских ежей практически отсутствует, за исключением немногочисленных видов с развитой заботой о потомстве (у них самки имеют специальные выводковые камеры для вынашивания молоди). Что же имели в виду авторы, включив морских ежей в число животных с документированным SSB? Дело в том, что для нескольких видов был показан рост вероятности вымётывания в воду половых продуктов в том случае, если поблизости есть другие морские ежи (это, очевидно, повышает шансы выпущенных в воду гамет найти себе партнеров). Но при этом пол соседей, как выяснилось, абсолютно не важен. Судя по всему, морские ежи как-то чуют сородичей, но не способны отличить самца от самки. Что ж, выметывание гамет в ответ на присутствие поблизости сородичей того же пола действительно можно трактовать как SSB.
По мнению авторов, именно такое, как у морских ежей, неразборчивое половое поведение, не предусматривающее выбора партнера по половому признаку, могло быть эволюционно исходным состоянием для раздельнополых животных — а вовсе не строгое DSB, как принято считать. В этом есть определенная логика, поскольку неразборчивое половое поведение явно проще избирательного. В этом случае животные не нуждаются ни в половом диморфизме (то есть в признаках, по которым можно определить пол потенциального партнера), ни в умении разбираться в этих тонкостях, ни в дополнительных рецепторах или органах чувств, которыми, вероятно, пришлось бы обзаводиться ради такого умения.
Таким образом, авторы считают, что постулат об эволюционной первичности строгого DSB, обычно принимаемый «по умолчанию», нуждается в пересмотре. Проще предположить, что исходно половое поведение животных было устроено примерно как у морских ежей: чуешь сородича — выметываешь половые продукты. В дальнейшем в разных эволюционных ветвях под действием отбора могли развиться разнообразные сложные (и порой весьма затратные) механизмы точной идентификации наиболее подходящих половых партнеров — если выигрыши от такого приобретения перевешивали затраты. Но принимать за аксиому первичность наличия таких механизмов нет веских оснований.
При неразборчивом половом поведении SSB и DSB, по идее, должны встречаться в популяции примерно с равной частотой. Но не слишком ли это расточительно? Ведь на однополое сексуальное поведение тратятся ресурсы организма (время, силы, гаметы), которые можно было бы использовать продуктивнее, направив их на DSB.
Принято считать, что в типичном случае SSB очень сильно снижает дарвиновскую приспособленность (репродуктивный успех) индивида именно из-за этих «непродуктивных» трат. Это еще один постулат (не всегда высказываемый в явном виде, но принимаемый как нечто самоочевидное), на котором обычно строятся теоретические объяснения распространенности SSB у тех или иных видов животных.
Эти объяснения обычно исходят из того, что в популяции со строгим и всеобщим DSB появляется мутация — новый аллель, повышающий вероятность SSB, и нужно понять, при каких условиях этот аллель будет поддержан отбором. Очевидно, что это произойдет лишь в том случае, если аллель даёт какие-то репродуктивные преимущества, перевешивающие приносимый им же репродуктивный ущерб. Если ущерб велик (а по умолчанию принято считать, что он велик), то и преимущества должны быть значительными. Они могут быть прямыми или косвенными. В первом случае SSB само по себе повышает шансы индивида на успешное размножение. Например, можно вообразить ситуацию, в которой самки охотнее спариваются с теми самцами, которые у них на глазах спаривались с кем-то еще — неважно, с самками или самцами (см.: Вид ухаживающего самца привлекает мексиканских рыбок независимо от его ориентации, «Элементы», 18.01.2013).
Во втором случае SSB дает какое-то важное преимущество окольным путем. Например, могут существовать аллели, повышающие репродуктивный успех самок, но приводящие — в качестве побочного эффекта — к росту вероятности SSB у самцов, что, как принято считать, сильно снижает репродуктивный успех самца (A. Camperio Ciani et al., 2008. Sexually Antagonistic Selection in Human Male Homosexuality). Такой аллель будет поддержан отбором только в том случае, если его положительное влияние на репродуктивный успех самок перевешивает его отрицательное влияние на репродуктивный успех самцов.
По мнению авторов, в подобных теоретических построениях, во-первых, необязательно начинать с популяции, в которой господствует DSB, а затем просчитывать судьбу вновь возникшей мутации, повысившей вероятность SSB. Может быть, правильнее начинать с «неразборчивого» состояния и думать о том, подхватит ли отбор мутацию, способствующую более точному выбору партнера — с учетом всех связанных с этим издержек.
Во-вторых, SSB вовсе не обязано сильно снижать репродуктивный успех. Речь, конечно, идет не об облигатном SSB. Понятно, что индивид, демонстрирующий половое поведение только по отношению к представителям своего пола, вряд ли вообще оставит потомство. Однако неразборчивое половое поведение, ведущее к эпизодическому SSB, во многих случаях, по идее, не должно быть таким уж затратным.
Есть немало примеров, это подтверждающих. Так, эволюционные эксперименты на жуках Tribolium castaneum (рис. 1, i) показали, что если доля самок в популяции высока, то самцы со временем полностью утрачивают половую избирательность и начинают спариваться с самцами так же охотно, как и с самками (см. статью Натальи Резник. Ошибочка вышла?). По-видимому, в этом случае «непродуктивный» расход ресурсов (включая и расход сперматофоров, которые у этих жуков выбрасываются при однополых спариваниях точно так же, как и при разнополых) не настолько обременителен, чтобы отбор продолжал поддерживать дорогостоящие механизмы безошибочного различения сородичей по полу.
В случае с морскими ежами тоже очевидно, что затратность SSB не слишком высока, — особенно если учесть, что скопления морских ежей обычно состоят не из двух, а из большего числа особей, так что среди них, скорее всего, найдутся и самцы, и самки. Поэтому у всякой выброшенной в воду гаметы будет ненулевой шанс найти себе пару, даже если члены группы не умеют определять пол соседей.
Кроме того, у многих животных наблюдается половое поведение, не связанное с зачатием, но выполняющее иные социальные функции — это тоже согласуется с идеей о том, что затратность «избыточного» сексуального поведения не обязательно должна быть высокой (сексуальные «игры» есть даже у пауков, см.: J. N. Pruitt, 2011. Non-conceptive sexual behavior in spiders: a form of play associated with body condition, personality type, and male intrasexual selection).
Нужно помнить и том, что чрезмерная разборчивость при выборе партнера сопряжена с риском упущенных репродуктивных возможностей. Никакая избирательность не может быть абсолютно безошибочной, но животные, которые попытаются довести ее до совершенства, чтобы исключить вероятность SSB, рискуют упустить свой шанс на размножение. Например, излишне разборчивый самец может упустить шанс спариться с самкой, которая почему-то показалась ему недостаточно женственной. Эти рассуждения согласуются с тем, что некоторые животные в ходе эволюции, по-видимому, научились подстраивать под ситуацию уровень своей половой избирательности. Например, самцы жуков-могильщиков становятся менее избирательными — и чаще демонстрируют SSB, — если в период своей жизни, предшествовавший размножению, они редко встречали самок, то есть как бы приучались к мысли, что самка — большая редкость и ее нельзя упускать; дополнительные эксперименты позволили исключить альтернативные интерпретации этого результата (K. C. Engel et al., 2015. Acceptance threshold theory can explain occurrence of homosexual behaviour).
Особенно далеко это зашло у некоторых бесхвостых амфибий. Так, самцы серой жабы (рис. 1, o) в период размножения пытаются спариться практически с любым движущимся объектом, примерно подходящим по размеру. Если этим объектом оказывается другой самец того же вида, он издает особые звуки, и его сразу отпускают. Возможно, этот условный сигнал является адаптацией, развившейся специально для минимизации ущерба от SSB. Лягушки, схваченные самцом жабы, тоже кричат по-своему, но они не знают жабьего «пароля», и поэтому им намного труднее освободиться от страстных объятий (A. Marco, M. Lizana, 2002. The absence of species and sex recognition during mate search by male common toads, Bufo bufo).
Отличие предлагаемого авторами нового взгляда на эволюцию SSB от традиционных представлений схематично показано на рис. 2.
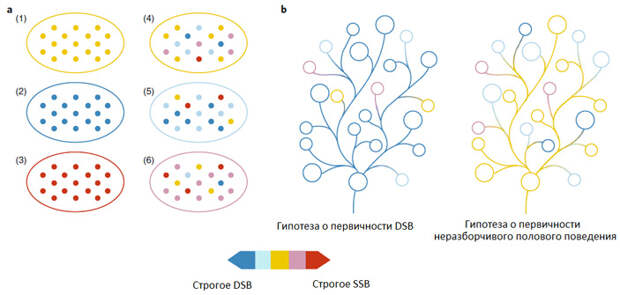
Рис. 2. Изменчивость и эволюция полового поведения. Левая схема (а) иллюстрирует простую мысль о том, что как особи, так и популяции могут различаться по соотношению SSB и DSB. Популяции могут быть мономорфными (1, 2, 3), то есть включать особей с одинаковыми наклонностями, и полиморфными (4, 5, 6). Мономорфная популяция, все особи в которой практикуют только SSB (3), нежизнеспособна, однако остальные варианты имеют право на существование. Справа (b) показаны эволюционные деревья, соответствующие традиционному представлению о первичности DSB и новой идее о первичности неразборчивого полового поведения. Изображение из обсуждаемой статьи в Nature Ecology & Evolution
Авторы подчеркивают, что их идея о первичности неразборчивого полового поведения и о том, что сдвиги в сторону преобладания DSB происходили независимо в разных ветвях (когда выигрыш от более точного выбора партнера почему-либо перевешивал связанные с этим затраты), может и должна быть проверена при помощи филогенетического анализа. Для этого нужно собрать как можно более точные и подробные данные о соотношении SSB и DSB у разных видов животных, посмотреть, как распределяется этот признак по концевым веточкам эволюционного дерева, оценить «филогенетический сигнал» и, наконец, реконструировать наиболее вероятное состояние признака в точках ветвления дерева (пример подобного исследования рассмотрен в новости Человеческая агрессивность имеет глубокие эволюционные корни, «Элементы», 03.10.2016).
Однако для такого анализа, по мнению авторов, пока еще недостаточно данных. Сведения о встречаемости SSB у разных видов крайне фрагментарны. Исследователи часто игнорируют наблюдаемые случаи SSB или не классифицируют их как половое поведение, а относят к другим категориям социальных взаимодействий. У многих животных половая принадлежность не определяется на глаз, и поэтому наблюдатели могут принимать SSB за DSB. Наконец, в лабораторных исследованиях половой избирательности животным редко дают возможность выбирать между представителями своего и противоположного пола. По мнению авторов, корректный филогенетический анализ станет возможен лишь после того, как исследователи начнут более внимательно и аккуратно отслеживать случаи SSB и перестанут считать такое поведение аномальным.
В заключительном разделе статьи авторы, к сожалению, не смогли удержаться от замечания, которое, на мой взгляд, смотрелось бы уместнее в газетной статье, чем в научной. Они отметили, что если бы любой другой признак встречался в самых разных группах животных так же часто, как SSB, то теоретики давно уже задумались бы о том, не является ли такое состояние эволюционно первичным (а не появившимся много раз независимо, каждый раз в силу каких-то особых причин). Авторы усматривают в этом влияние традиционной европейской патриархальной культуры, в рамках которой DSB считалось единственным социально приемлемым вариантом полового поведения. С отказом от устаревших культурных норм открывается, по мнению авторов, возможность для более беспристрастного изучения эволюции полового поведения.
Источник: Julia D. Monk, Erin Giglio, Ambika Kamath, Max R. Lambert, Caitlin E. McDonough. An alternative hypothesis for the evolution of same-sex sexual behaviour in animals // Nature Ecology & Evolution. 2019. V. 3. P. 1622–1631. DOI: 10.1038/s41559-019-1019-7.
См. также:
1) Марина Бутовская. Гомосексуализм и эволюция. Издательство «Век 2», 2006.
2) Наталья Резник. Ошибочка вышла? // «Троицкий вариант», №10, 2018.
3) Вид ухаживающего самца привлекает мексиканских рыбок независимо от его ориентации, «Элементы», 18.01.2013.
4) Доминантные самцы оказываются в проигрыше, когда их слишком много, «Элементы», 21.11.2011.
Свежие комментарии